Клетки крови человека функции. Клетки крови человека
Любые изменения состава крови у человека имеют высокую диагностическую ценность для установления причины заболевания и идентификации возбудителя.
Кровь, по сути своей является суспензией, которая подразделяется на жидкую плазму и форменные элементы. В среднем, составляющие крови на 40% состоит их элементов, распределённых в плазме. Форменные элементы на 99% состоят из эритроцитов (ἐρυθρός — красный). Процентное отношение объёма (RBC) к общей ёмкости крови называют HCT (гематокрит). При потере кровью внушительного объёма жидкости, говорят о . Такое состояние наступает, когда процент плазмы опускается ниже 55%.
Причинами патологии крови могут быть:
- Понос;
- Рвота;
- Ожоговая болезнь;
- Обезвоживание организма при тяжёлой работе, в результате спортивных состязаний и длительного пребывания на жаре.
По особенностям отзыва лейкоцитов на происходящие изменения, делают вывод о наличии инфекции и её разновидности, определяют этапы патологического процесса, восприимчивость организма к назначенному лечению. Изучение лейкоформулы позволяет обнаруживать опухолевые патологии. При детальной расшифровке лейкоцитарной формулы, можно установить не только наличие лейкоза или лейкопении, но уточнить, каким видом онкологии человек страдает.
Немаловажное значение имеет обнаружение повышенного вброса в периферийную кровь клеток-предшественников лейкоцитов. Это говорит об извращении синтеза лейкоцитов, приводящего к онкологии крови.
У человека (PLT) — это мелкие клетки, лишённые ядра, задачей которых является сохранение целостности кровяного русла. PLT способны слипаться, приклеиваться к разнообразным поверхностям, образуя тромбы при разрушениях стенок сосудов. Тромбоциты в крови содействуют лейкоцитам в ликвидации чужеродных агентов, увеличивая просвет капилляров.
В организме ребёнка кровь занимает до 9% массы тела. У взрослого процент самой главной соединительной ткани организма падает до семи, что составляет, не менее пяти литров.
Соотношение упомянутых выше компонентов крови может меняться по причине болезни, либо, как следствие иных обстоятельств.

Причины изменения состава крови у взрослого и ребенка могут стать:
- Несбалансированное питание;
- Возраст;
- Физиологические состояния;
- Климат;
- Вредные привычки.
Чрезмерное употребление жиров провоцирует кристаллизацию холестерина на стенках сосудов. Избыток белков, из-за увлечения мясными продуктами выводится из организма в виде мочевой кислоты. Неумеренное потребление кофе провоцирует эритроцитоз, гипергликемию и и состав крови человека меняется.
Дисбаланс поступления с пищей или усвоения железа, фолиевой кислоты и цианкобаламина приводит к падению гемоглобина. Голодание является причиной роста билирубина.
Мужчины, образ жизни которых предполагает более высокие физические напряжения, по сравнению с женщинами, нуждаются в большем количестве кислорода, что проявляется в повышении числа RВС и концентрации гемоглобина.
Нагрузки на организм пожилых постепенно уменьшаются, уводя вниз показатели крови.
Горцы, постоянно находящиеся в условиях нехватки кислорода компенсируют её повышением уровня RВС и НВ. Выведение из организма курильщика повышенного количества шлаков и токсинов сопровождается лейкоцитозом.
Оптимизировать показатели крови можно во время болезни. Первым делом, нужно наладить полноценное питание. Избавиться от вредных привычек. Ограничить употребление кофе, бороться с адинамией посредством умеренной физической нагрузки. Кровь отблагодарит хозяина, готового бороться за сохранение здоровья. Вот так вот выглядит состав крови человека если разбирать его по ее компонентам.
Кровь, беспрерывно циркулирующая в замкнутой системе кровеносных сосудов, выполняет в организме важнейшие функции: транспортную, дыхательную, регуляторную и защитную. Она обеспечивает относительное постоянство внутренней среды организма.
Кровь — это разновидность соединительной ткани, состоящей из жидкого межклеточного вещества сложного состава — плазмы н взвешенных в ней клеток — форменных элементов крови: эритроцитов (красных кровяных клеток), лейкоцитов (белых кровяных клеток) и тромбоцитов (кровяных пластинок). В 1 мм 3 крови содержится 4,5–5 млн. эритроцитов, 5–8 тыс. лейкоцитов, 200–400 тыс. тромбоцитов.
В организме человека количество крови составляет в среднем 4,5–5 л или 1/13 массы его тела. Плазма крови по объему составляет 55–60%, а форменные элементы 40–45%. Плазма крови представляет собой желтоватую полупрозрачную жидкость. В ее состав входит вода (90–92%), минеральные и органические вещества (8–10%), 7% белков. 0,7% жиров, 0.1% — глюкозы, остальная часть плотного остатка плазмы — гормоны, витамины, аминокислоты, продукты обмена веществ.
Форменные элементы крови
Эритроциты — безъядерные красные кровяные клетки, имеющие форму двояковогнутых дисков. Такая форма увеличивает поверхность клетки в 1.5 раза. Цитоплазма эритроцитов содержит белок гемоглобин — сложное органическое соединение, состоящее из белка глобина и пигмента крови гема, в состав которого входит железо.
Основная функция эритроцитов — транспортировка кислорода и углекислого газа. Эритроциты развиваются из ядерных клеток в красном костном мозге губчатого вещества кости. В процессе созревания они теряют
Клетка — Википедия


Кле́тка — структурно-функциональная элементарная единица строения и жизнедеятельности всех организмов (кроме вирусов и вироидов — форм жизни, не имеющих клеточного строения). Обладает собственным обменом веществ, способна к самовоспроизведению. Организм, состоящий из одной клетки, называется одноклеточным (многие простейшие и бактерии). Раздел биологии, занимающийся изучением строения и жизнедеятельности клеток, называется цитологией. Также принято говорить о биологии клетки, или клеточной биологии.
 Срез пробкового дерева из книги Роберта Гука «Микрография», 1635—1703 Первое наблюдение за клетками, с использованием раннего микроскопа[1]. Это привело к развитию теории клеток.
Срез пробкового дерева из книги Роберта Гука «Микрография», 1635—1703 Первое наблюдение за клетками, с использованием раннего микроскопа[1]. Это привело к развитию теории клеток.Первым человеком, увидевшим клетки, был английский учёный Роберт Гук (известный открытием закона Гука). В 1665 году, пытаясь понять, почему пробковое дерево хорошо плавает, Гук стал рассматривать тонкие срезы пробки с помощью усовершенствованного им микроскопа. Он обнаружил, что пробка разделена на множество крошечных ячеек, напомнивших ему соты в ульях медоносных пчёл, и он назвал эти ячейки клетками (по-английски cell означает «ячейка, клетка»).
В 1675 году итальянский врач Марчелло Мальпиги подтвердил клеточное строение растений, а в 1681 году — английский ботаник Неемия Грю. О клетке стали говорить как о «пузырьке, наполненном питательным соком». В 1674 году голландский мастер Антоний ван Левенгук с помощью микроскопа впервые увидел в капле воды «зверьков» — движущиеся живые организмы (инфузории, амёбы, бактерии). Также Левенгук впервые наблюдал животные клетки — эритроциты и сперматозоиды. Таким образом, к началу XVIII века учёные знали, что под большим увеличением растения имеют ячеистое строение, и видели некоторые организмы, которые позже получили название одноклеточных. В 1802—1808 годах французский исследователь Шарль-Франсуа Мирбель установил, что растения состоят из тканей, образованных клетками. Ж. Б. Ламарк в 1809 году распространил идею Мирбеля о клеточном строении и на животные организмы. В 1825 году чешский учёный Я. Пуркине открыл ядро яйцеклетки птиц, а в 1839 ввёл термин «протоплазма». В 1831 году английский ботаник Р. Броун впервые описал ядро растительной клетки, а в 1833 году установил, что ядро является обязательным органоидом клетки растения. С тех пор главным в организации клеток считается не оболочка, а содержимое.
Клеточная теория[править | править код]
Клеточная теория строения организмов была сформирована в 1839 году немецкими учёными, зоологом Теодором Шванном и ботаником М. Шлейденом, и включала в себя три положения. В 1858 году Рудольф Вирхов дополнил её ещё одним положением, однако в его идеях присутствовал ряд ошибок: так, он предполагал, что клетки слабо связаны друг с другом и существуют каждая «сама по себе». Лишь позднее удалось доказать целостность клеточной системы.
В 1878 году русским учёным И. Д. Чистяковым открыт митоз в растительных клетках; в 1878 году В. Флемминг и П. И. Перемежко обнаруживают митоз у животных. В 1882 году В. Флемминг наблюдает мейоз у животных клеток, а в 1888 году Э. Страсбургер — у растительных.
Клеточная теория является одной из основополагающих идей современной биологии, она стала неопровержимым доказательством единства всего живого и фундаментом для развития таких дисциплин, как эмбриология, гистология и физиология. На сегодняшний день теория содержит такие утверждения:
- Клетка — элементарная единица строения, функционирования, размножения и развития всех живых организмов. Вне клетки нет жизни.
- Клетка — целостная система, содержащая большое количество связанных друг с другом элементов — органелл.
- Клетки различных организмов похожи (гомологичны) по строению и основным свойствам и имеют общее происхождение.
- Увеличение количества клеток происходит путём их деления, после репликации их ДНК: клетка — от клетки.
- Многоклеточный организм — система из большого количества клеток, объединённых в системы тканей и органов, связанных между собой гуморальной и нервной регуляциями.
- Клетки многоклеточных организмов обладают одинаковым полным фондом генетического материала этого организма, всеми возможными потенциями для проявления этого материала, — но отличаются по уровню экспрессии (работы) отдельных генов, что приводит к их морфологическому и функциональному разнообразию — дифференцировке[2].
Количество и формулировки отдельных положений современной клеточной теории в разных источниках могут отличаться.
Впервые клетки удалось увидеть только после создания оптических (световых) микроскопов. С того времени микроскопия остается одним из важнейших методов исследования клеток. Световая микроскопия, несмотря на небольшое разрешение, позволяла наблюдать за живыми клетками. В XX веке была изобретена электронная микроскопия, которая позволила изучить ультраструктуру клеток.
Для изучения функций клеток и их частей используют разнообразные биохимические методы — как препаративные, например фракционирование методом дифференциального центрифугирования, так и аналитические. Для экспериментальных и практических целей используют методы . Все упомянутые методические подходы могут использоваться в сочетании с методами культуры клеток.
Оптическая микроскопия[править | править код]
В оптическом микроскопе увеличение объекта достигается благодаря серии линз, через которые проходит свет. Максимальное увеличение составляет более 1000 раз. Также важной характеристикой является разрешение — расстояние между двумя точками, которые ещё распознаются отдельно. Разрешение характеризует чёткость изображения. Эта величина ограничивается длиной световой волны, и даже при использовании самого коротковолнового света — ультрафиолетового — можно достичь разрешения только около 200 нм; такое разрешение было получено ещё в конце XIX века. Малейшие структуры, которые можно наблюдать под оптическим микроскопом, это митохондрии и бактерии. Их линейный размер составляет примерно 500 нм. Однако объекты размером меньше 200 нм видны в световом микроскопе, если они сами излучают свет. Эта особенность используется в флуоресцентной микроскопии, когда клеточные структуры или отдельные белки связываются со специальными флуоресцентными белками или антителами с флуоресцентными метками. На качество изображения, полученного с помощью оптического микроскопа, влияет также контрастность — её можно увеличить, используя различные методы окраски клеток. Для изучения живых клеток используют фазово-контрастную, дифференциальную интерференционно-контрастную и темнопольную микроскопию. Конфокальные микроскопы позволяют улучшить качество флуоресцентных изображений[3][4].
Изображения, полученные с помощью оптической микроскопииЭлектронная микроскопия[править | править код]
В 30-х годах XX века был сконструирован электронный микроскоп, в котором вместо света через объект пропускается пучок электронов. Теоретический предел разрешения для современных электронных микроскопов составляет около 0,002 нм, однако из практических причин для биологических объектов достигается разрешение только около 2 нм. С помощью электронного микроскопа можно изучать ультраструктуру клеток. Различают два основных типа электронной микроскопии: сканирующую и трансмиссионную. Сканирующая (растровая) электронная микроскопия (РЭМ) используется для изучения поверхности объекта. Образцы зачастую покрывают тонкой плёнкой золота. РЭМ позволяет получать объёмные изображения. Трансмиссионная (просвечивающая) электронная микроскопия (ПЭМ) — используется для изучения внутреннего строения клетки. Пучок электронов пропускается через объект, предварительно обработанный тяжёлыми металлами, которые накапливаются в определённых структурах, увеличивая их электронную плотность. Электроны рассеиваются на участках клетки с большей электронной плотностью, в результате чего на изображениях эти области выглядят темнее[3][4].
Фракционирование клеток[править | править код]
Для установления функций отдельных компонентов клетки важно выделить их в чистом виде, чаще всего это делается с помощью метода дифференциального центрифугирования. Разработаны методики, позволяющие получить чистые фракции любых клеточных органелл. Получение фракций начинается с разрушения плазмалеммы и образования клеток. Гомогенат последовательно центрифугируется при различных скоростях, на первом этапе можно получить четыре фракции: (1) ядер и крупных обломков клеток, (2) митохондрий, пластид, лизосом и пероксисом, (3) — пузырьков аппарата Гольджи и эндоплазматического ретикулума, (4) рибосом, в останутся белки и более мелкие молекулы. Дальнейшее дифференциальное центрифугирование каждой из смешанных фракций позволяет получить чистые препараты органелл, к которым можно применять разнообразные биохимические и микроскопические методы[2].
Все клеточные формы жизни на Земле можно разделить на два надцарства на основании строения составляющих их клеток:
- прокариоты (доядерные) — более простые по строению, возникли в процессе эволюции раньше;
- эукариоты (ядерные) — более сложные, возникли позже. Клетки, составляющие тело человека, являются эукариотическими.
Несмотря на многообразие форм, организация клеток всех живых организмов подчинена единым структурным принципам.
Содержимое клетки отделено от окружающей среды плазматической мембраной, или плазмалеммой. Внутри клетка заполнена цитоплазмой, в которой расположены различные органеллы и клеточные включения, а также генетический материал в виде молекулы ДНК. Каждая из органелл клетки выполняет свою особую функцию, а в совокупности все они определяют жизнедеятельность клетки в целом.
Прокариотическая клетка[править | править код]
Прокариоты (от др.-греч. πρό — ‘перед’, ‘до’ и κάρῠον — ‘ядро’) — организмы, не обладающие, в отличие от эукариот, оформленным клеточным ядром и другими внутренними мембранными органоидами (за исключением плоских цистерн у фотосинтезирующих видов, например, у цианобактерий). Единственная крупная кольцевая (у некоторых видов — линейная) двухцепочечная молекула ДНК, в которой содержится основная часть генетического материала клетки (так называемый нуклеоид) не образует комплекса с белками-гистонами (так называемого хроматина). К прокариотам относятся бактерии, в том числе цианобактерии (сине-зелёные водоросли), и археи. Потомками прокариотических клеток являются органеллы эукариотических клеток — митохондрии и пластиды. Основное содержимое клетки, заполняющее весь её объём, — вязкая зернистая цитоплазма.
Эукариотическая клетка[править | править код]
Эукариоты (эвкариоты, от др.-греч. εὖ ‘хорошо’, ‘полностью’ и κάρῠον — ‘ядро’) — организмы, обладающие, в отличие от прокариот, оформленным клеточным ядром, отграниченным от цитоплазмы ядерной оболочкой. Генетический материал заключён в нескольких линейных двухцепочных молекулах ДНК (в зависимости от вида организмов их число на ядро может колебаться от двух до нескольких сотен), прикреплённых изнутри к мембране клеточного ядра и образующих у подавляющего большинства (кроме динофлагеллят) комплекс с белками-гистонами, называемый хроматином. В клетках эукариот есть система внутренних мембран, образующих, помимо ядра, ряд других органоидов (эндоплазматическая сеть, аппарат Гольджи и др.). Кроме того, у подавляющего большинства имеются постоянные внутриклеточные симбионты-прокариоты — митохондрии, а у водорослей и растений — также и пластиды.

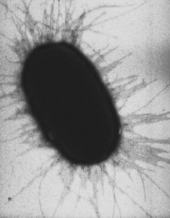 Фимбрии кишечной палочки, которые позволяют ей прикрепляться к субстрату (ОМ)
Фимбрии кишечной палочки, которые позволяют ей прикрепляться к субстрату (ОМ) 
Клетки двух основных групп прокариот — бактерий и архей — похожи по структуре, характерными их признаками являются отсутствие ядра и мембранных органелл.
Основными компонентами прокариотической клетки являются:
- Клеточная стенка, которая окружает клетку извне, защищает её, придаёт устойчивую форму, предотвращающую от осмотического разрушения. У бактерий клеточная стенка состоит из пептидогликана (муреина), построенного из длинных полисахаридных цепей, соединённых между собой короткими пептидными перемычками. По строению клеточной стенки различают две группы бактерий:
- Клеточная стенка архей не содержит муреина, а построена в основном из разнообразных белков и полисахаридов[4].
- Капсула — имеющаяся у некоторых бактерий слизистая оболочка, расположенная снаружи от клеточной стенки. Состоит в основном из разнообразных белков, углеводов и уроновых кислот. Капсулы защищают клетки от высыхания, могут помогать бактериям в колониях удерживаться вместе, а индивидуальным бактериям — прикрепляться к различным субстратам. Кроме этого, капсулы предоставляют клетке дополнительную защиту: например, капсулированные штаммы пневмококков свободно размножаются в организме и вызывают воспаление лёгких, тогда как некапсулированные быстро уничтожаются иммунной системой и являются абсолютно безвредными[5].
- Пили или ворсинки — тонкие волоскоподобные выросты, что присутствуют на поверхности бактериальных клеток. Существуют различные типы пилей, из которых наиболее распространёнными являются:
- Жгутики — органеллы движения некоторых бактерий. Бактериальный жгутик построен значительно проще эукариотического, и он в 10 раз тоньше, не покрыт плазматической мембраной и состоит из одинаковых молекул белков, которые образуют цилиндр. В мембране жгутик закреплён при помощи базального тела[4].
- Плазматическая и внутренние мембраны. Общий принцип устройства клеточных мембран не отличается от эукариот, однако в химическом составе мембраны есть немало различий, в частности, в мембранах прокариот отсутствуют молекулы холестерина и некоторых липидов, присущих мембранам эукариот. Большинство прокариотических клеток (в отличие от эукариотических) не имеют внутренних мембран, которые разделяют цитоплазму на отдельные компартменты. Только у некоторых фотосинтетических и аэробных бактерий плазмалемма образует вгибание внутрь клетки, что выполняет соответствующие метаболические функции[4].
- Нуклеоид — не ограниченный мембранами участок цитоплазмы, в котором расположена кольцевая молекула ДНК — «бактериальная хромосома», где хранится весь генетический материал клетки[4].
- Плазмиды — небольшие дополнительные кольцевые молекулы ДНК, несущие обычно всего несколько генов. Плазмиды, в отличие от бактериальной хромосомы, не являются обязательным компонентом клетки. Обычно они придают бактерии определённые полезные для неё свойства, такие как устойчивость к антибиотикам, способность усваивать из среды определённые энергетические субстраты, способность инициировать половой процесс и т. д.[4][5]
- Рибосомы прокариот, как и у всех других живых организмов, отвечают за осуществление процесса трансляции (одного из этапов биосинтеза белка). Однако бактериальные рибосомы несколько меньше, чем эукариотические (коэффициенты седиментации 70S и 80S соответственно), и имеют другой состав белков и РНК. Из-за этого бактерии, в отличие от эукариот, чувствительны к таким антибиотикам, как эритромицин и тетрациклин, которые избирательно действуют на 70S-рибосомы[5].
- Эндоспоры — окружённые плотной оболочкой структуры, содержащие ДНК бактерии и обеспечивающее выживание в неблагоприятных условиях. К образованию эндоспор способны лишь некоторые виды прокариот, например представители родов Clostridium (C. tetani — возбудитель столбняка, C. botulinum — возбудитель ботулизма, C. perfringens — возбудитель газовой гангрены и т. п.) и Bacillus (в частности B. anthracis — возбудитель сибирской язвы). Для образования эндоспоры клетка реплицирует свою ДНК и окружает копию плотной оболочкой, из созданной структуры удаляется избыток воды, и в ней замедляется метаболизм[4]. Споры бактерий могут выдерживать довольно жёсткие условия среды, такие как длительное высушивание, кипячение, коротковолновое облучение и др.[5]
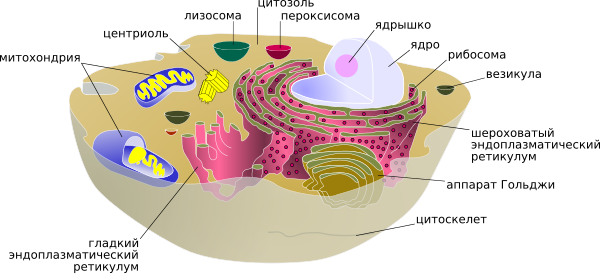 Схематическое изображение животной клетки (подписи составных частей — ссылки на статьи про них).
Схематическое изображение животной клетки (подписи составных частей — ссылки на статьи про них).Поверхностный комплекс животной клетки[править | править код]
Состоит из гликокаликса, плазмалеммы и расположенного под ней кортикального слоя цитоплазмы. Плазматическая мембрана называется также плазмалеммой, наружной клеточной мембраной, цитолеммой и т. д. Её толщина — около 10 нанометров. Обеспечивает разграничение клетки и внешней среды, а также пропускание внутрь и наружу некоторых веществ.
На сохранение целостности своей мембраны клетка не тратит энергии: молекулы удерживаются по тому же принципу, по которому удерживаются вместе молекулы жира — гидрофобным частям молекул термодинамически выгоднее располагаться в непосредственной близости друг к другу. Гликокаликс представляет собой «заякоренные» в плазмалемме молекулы олигосахаридов, полисахаридов, гликопротеинов и гликолипидов. Гликокаликс выполняет рецепторную и маркерную функции.
Плазматическая мембрана животных клеток в основном состоит из фосфолипидов и липопротеидов со вкрапленными в неё молекулами белков, в частности, поверхностных антигенов и рецепторов.
В кортикальном (прилегающем к плазматической мембране) слое цитоплазмы находятся специфические элементы цитоскелета — упорядоченные определённым образом актиновые микрофиламенты. Основной и самой важной функцией кортикального слоя (кортекса) являются псевдоподиальные реакции: выбрасывание, прикрепление и сокращение псевдоподий. При этом микрофиламенты перестраиваются, удлиняются или укорачиваются. От структуры цитоскелета кортикального слоя зависит также форма клетки (например, наличие микроворсинок).
Структура цитоплазмы[править | править код]
Жидкую составляющую цитоплазмы также называют цитозолем. Под световым микроскопом казалось, что клетка заполнена чем-то вроде жидкой плазмы или золя, в котором «плавают» ядро и другие органеллы. На самом деле это не так. Внутреннее пространство эукариотической клетки строго упорядочено. Передвижение органоидов координируется при помощи специализированных транспортных систем, так называемых микротрубочек, служащих внутриклеточными «дорогами», и специальных белков динеинов и кинезинов, играющих роль «двигателей». Отдельные белковые молекулы также не диффундируют свободно по всему внутриклеточному пространству, а направляются в необходимые компартменты при помощи специальных сигналов на их поверхности, узнаваемых транспортными системами клетки.
Рибосомы[править | править код]
Рибосомы — органоиды, необходимые клетке для синтеза белка. Их размер составляет примерно 20—30 нм. В клетке их насчитывается несколько миллионов. Рибосомы образованы из двух субъединиц: большой и малой, состоящих из четырёх молекул РНК и нескольких молекул белков. У эукариотических клеток рибосомы встречаются не только в цитоплазме, но и в митохондриях и хлоропластах. Рибосомы формируются в области ядрышек, а затем через ядерные поры выходят в цитоплазму.
Эндоплазматический ретикулум[править | править код]
В эукариотической клетке существует система переходящих друг в друга мембранных отсеков (трубочек, мешочков и плоских цистерн разных размеров), которая называется эндоплазматическим ретикулумом (или эндоплазматическая сеть, ЭПР или ЭПС). Ту часть ЭПР, к мембранам которого прикреплены рибосомы, относят к гранулярному (или шероховатому) эндоплазматическому ретикулуму, на его мембранах происходит синтез белков. Те компартменты, на стенках которых нет рибосом, относят к агранулярному (или гладкому) ЭПР, принимающему участие в синтезе липидов. Внутренние пространства гладкого и гранулярного ЭПС не изолированы, а переходят друг в друга и сообщаются с просветом ядерной оболочки.
Аппарат Гольджи[править | править код]
Аппарат Гольджи представляет собой стопку плоских мембранных цистерн, несколько расширенных ближе к краям. В цистернах аппарата Гольджи созревают некоторые белки, синтезированные на мембранах гранулярного ЭПР и предназначенные для секреции или образования лизосом. Аппарат Гольджи асимметричен — цистерны, располагающиеся ближе к ядру клетки (цис-Гольджи) содержат наименее зрелые белки, к этим цистернам непрерывно присоединяются мембранные пузырьки — везикулы, отпочковывающиеся от эндоплазматического ретикулума. По-видимому, при помощи таких же пузырьков происходит дальнейшее перемещение созревающих белков от одной цистерны к другой. В конце концов от противоположного конца органеллы (транс-Гольджи) отпочковываются пузырьки, содержащие полностью зрелые белки.
Ядро[править | править код]
Клеточное ядро содержит молекулы ДНК, на которых записана генетическая информация организма. В ядре происходит репликация — удвоение молекул ДНК, а также транскрипция — синтез молекул РНК на матрице ДНК. В ядре же синтезированные молекулы РНК претерпевают некоторые модификации (например, в процессе сплайсинга из молекул матричной РНК исключаются незначащие, бессмысленные участки), после чего выходят в цитоплазму. Сборка рибосом также происходит в ядре, в специальных образованиях, называемых ядрышками.
Компартмент для ядра — кариотека — образован за счёт расширения и слияния друг с другом цистерн эндоплазматической сети таким образом, что у ядра образовались двойные стенки за счёт окружающих его узких компартментов ядерной оболочки. Полость ядерной оболочки называется люменом или перинуклеарным пространством. Внутренняя поверхность ядерной оболочки подстилается ядерной ламиной, жесткой белковой структурой, образованной белками-ламинами, к которой прикреплены нити хромосомной ДНК. В некоторых местах внутренняя и внешняя мембраны ядерной оболочки сливаются и образуют так называемые ядерные поры, через которые происходит материальный обмен между ядром и цитоплазмой.
Лизосомы[править | править код]
Лизосома — небольшое тельце, ограниченное от цитоплазмы одинарной мембраной и имеющее вид пузырька. В ней находятся литические ферменты, способные расщепить практически любые природные органические соединения. Основная функция — автолиз — то есть расщепление отдельных органоидов, участков цитоплазмы клетки.
Цитоскелет[править | править код]
К элементам цитоскелета относят белковые фибриллярные структуры, расположенные в цитоплазме клетки: микротрубочки, актиновые и промежуточные филаменты. Микротрубочки принимают участие в транспорте органелл, входят в состав органоидов движения — ресничек и жгутиков, характерных для некоторых клеток (например, инфузорий, сперматозоидов), из микротрубочек строится митотическое веретено деления. Актиновые филаменты необходимы для поддержания формы клетки, псевдоподиальных реакций. Роль промежуточных филаментов, по-видимому, также заключается в поддержании структуры клетки. Белки цитоскелета составляют несколько десятков процентов от массы клеточного белка.
Центриоли[править | править код]
Центриоли представляют собой цилиндрические белковые структуры, расположенные вблизи ядра клеток животных (у растений центриолей нет, за исключением низших водорослей). Центриоль представляет собой цилиндр, боковая поверхность которого образована девятью наборами микротрубочек. Количество микротрубочек в наборе может колебаться для разных организмов от 1 до 3.
Вокруг центриолей находится так называемый центр организации цитоскелета, район, в котором группируются минус-концы микротрубочек клетки.
Перед делением клетка содержит две центриоли, расположенные под прямым углом друг к другу. В ходе митоза они расходятся к разным концам клетки, формируя полюса веретена деления. После цитокинеза каждая дочерняя клетка получает по одной центриоли, которая удваивается к следующему делению. Удвоение центриолей происходит не делением, а путём синтеза новой структуры, перпендикулярной существующей.
Центриоли, по-видимому, гомологичны базальным телам жгутиков и ресничек.
Митохондрии[править | править код]
Митохондрии — особые органеллы клетки, основной функцией которых является синтез АТФ — универсального носителя энергии. Дыхание (поглощение кислорода и выделение углекислого газа) происходит также за счёт энзиматических систем митохондрий.
Внутренний просвет митохондрий, называемый матриксом, отграничен от цитоплазмы двумя мембранами, наружной и внутренней, между которыми располагается межмембранное пространство. Внутренняя мембрана митохондрии образует складки — кристы, на которых размещаются ферменты, ускоряющие реакции окисления жиров и углеводов. В матриксе содержатся различные ферменты, принимающие участие в дыхании и синтезе АТФ. Центральное значение для синтеза АТФ имеет водородный потенциал внутренней мембраны митохондрии.
Митохондрии имеют свой со
строение, форма и функции. Особенности строения эритроцитов :: SYL.ru
Эритроцит, строение и функции которого мы рассмотрим в нашей статье, является важнейшей составляющей крови. Именно эти клетки осуществляют газообмен, обеспечивая дыхание на клеточном и тканевом уровне.
Эритроцит: строение и функции
Кровеносная система человека и млекопитающих животных характеризуется наиболее совершенным строением по сравнению с другими организмами. Она состоит из четырехкамерного сердца и замкнутой системы сосудов, по которым непрерывно циркулирует кровь. Эта ткань состоит из жидкой составляющей — плазмы, и ряда клеток: эритроцитов, лейкоцитов и тромбоцитов. Каждая клетка играет свою роль. Строение эритроцита человека обусловлено выполняемыми функциями. Это касается размера, формы и количества данных клеток крови.

Особенности строения эритроцитов
Эритроциты имеют форму двояковогнутого диска. Они не способны самостоятельно передвигаться в кровяном русле, подобно лейкоцитам. К тканям и внутренним органам они поступают благодаря работе сердца. Эритроциты — прокариотические клетки. Это означает, что они не содержат оформленного ядра. Иначе они не могли бы переносить кислород и углекислый газ. Эта функция выполняется благодаря наличию внутри клеток особого вещества — гемоглобина, который также определяет красный цвет крови человека.

Строение гемоглобина
Строение и функции эритроцитов во многом обусловлены особенностями именно этого вещества. В гемоглобин входят две составляющие. Это железосодержащий компонент, который называется гем, и белок глобин. Впервые расшифровать пространственную структуру этого химического соединения удалось английскому биохимику Максу Фердинанду Перуцу. За это открытие в 1962 году он был удостоен Нобелевской премии. Гемоглобин является представителем группы хромопротеинов. К ним относятся сложные белки, состоящие из простого биополимера и простетической группы. Для гемоглобина этой группой является гем. К данной группе относится и хлорофилл растений, который обеспечивает протекание процесса фотосинтеза.

Как происходит газообмен
У человека и других хордовых животных гемоглобин находится внутри эритроцитов, а у беспозвоночных растворен прямо в плазме крови. В любом случае химический состав этого сложного белка позволяет образовывать нестойкие соединения с кислородом и углекислым газом. Кровь, насыщенная кислородом, называется артериальной. Она обогащается данным газом в легких.
Из аорты она направляется в артерии, а потом — в капилляры. Эти самые мелкие сосуды подходят к каждой клетке организма. Здесь эритроциты отдают кислород и присоединяют основной продукт дыхания — углекислый газ. С током крови, которая уже является венозной, они поступают снова в легкие. В этих органах газообмен происходит в мельчайших пузырьках — альвеолах. Здесь гемоглобин отсоединяет углекислый газ, который удаляется из организма посредством выдоха, и кровь снова насыщается кислородом.
Такие химические реакции обусловлены наличием двухвалентного железа в геме. В результате соединения и разложения последовательно формируется окси- и карбгемоглобин. Но сложный белок эритроцитов может образовывать и стойкие соединения. К примеру, при неполном сгорании топлива выделяется угарный газ, который формирует с гемоглобином карбоксигемоглобин. Этот процесс ведет к гибели эритроцитов и отравлению организма, которое может привести к летальному исходу.

Что такое малокровие
Одышка, ощутимая слабость, шум в ушах, заметная бледность кожных покровов и слизистых оболочек может свидетельствовать о недостаточном количестве гемоглобина крови. Норма его содержания колеблется в зависимости от пола. У женщин этот показатель составляет 120 — 140 г на 1000 мл крови, а у мужчин достигает 180 г/л. Содержание гемоглобина в крови новорожденных детей самое большое. Оно превышает эту цифру у взрослых людей, достигая 210 г/л.
Недостаток гемоглобина является серьезным заболеванием, которое называется малокровием или анемией. Оно может быть вызвано недостатком в продуктах питания витаминов и солей железа, пристрастием к употреблению алкоголя, влиянием на организм радиационного загрязнения и других негативных экологических факторов.
Снижение количества гемоглобина может быть обусловлено и естественными факторами. К примеру, у женщин причиной анемии могут быть менструальный цикл или беременность. Впоследствии количество гемоглобина нормализируется. Временное снижение данного показателя наблюдается и у активных доноров, которые часто сдают кровь. Но повышенное количество эритроцитов также достаточно опасно и нежелательно для организма. Оно приводит к увеличению густоты крови и образованию тромбов. Часто повышение этого показателя наблюдается у людей, проживающих в высокогорных районах.
Нормализовать уровень гемоглобина возможно употребляя продукты питания, содержащие железо. К ним относятся печень, язык, мясо крупного рогатого скота, кролика, рыба, черная и красная икра. Продукты растительного происхождения также содержат необходимый микроэлемент, однако находящееся в них железо усваивается гораздо сложнее. К ним относятся плоды бобовых, гречневая крупа, яблоки, патока, красный перец и зелень.
Форма и размер
Строение эритроцитов крови характеризуется прежде всего их формой, которая достаточно необычна. Она действительно напоминает диск, вогнутый с двух сторон. Такая форма красных кровяных клеток не случайна. Она увеличивает поверхность эритроцитов и обеспечивает наиболее эффективное проникновение в них кислорода. Такая необычная форма способствует и увеличению количества данных клеток. Так, в норме в 1 кубическом мм крови человека содержится около 5 млн. эритроцитов, что также способствует наилучшему газообмену.

Строение эритроцитов лягушки
Ученые давно установили, что красные кровяные клетки человека обладают чертами строения, которые обеспечивают наиболее эффективный газообмен. Это касается и формы, и количества, и внутреннего содержимого. Это особенно очевидно, когда сравнивают строение эритроцитов крови человека и лягушки. У последних красные кровяные клетки имеют овальную форму и содержат ядро. Это значительно уменьшает содержание дыхательных пигментов. Эритроциты лягушки значительно крупнее человеческих, поэтому и концентрация их не так высока. Для сравнения: если у человека в кубическом мм их более 5 млн., то у земноводных эта цифра достигает 0,38.
Эволюция эритроцитов
Строение эритроцитов человека и лягушки позволяет сделать выводы об эволюционных преобразованиях подобных структур. Дыхательные пигменты встречаются еще у простейших инфузорий. В крови беспозвоночных они содержатся прямо в плазме. Но это значительно увеличивает густоту крови, что может привести к формированию тромбов внутри сосудов. Поэтому с течением времени эволюционные преобразования шли в сторону появления специализированных клеток, формирования их двояковогнутой формы, исчезновения ядра, уменьшения их размера и повышения концентрации.
Онтогенез красных кровяных клеток
Эритроцит, строение которого имеет ряд характерных особенностей, сохраняет жизнеспособность в течение 120 дней. В дальнейшем следует их разрушение в печени и селезенке. Главным кроветворным органом человека является красный костный мозг. В нем непрерывно происходит формирование новых эритроцитов из стволовых клеток. Первоначально они содержат ядро, которое по мере созревания разрушается и заменяется гемоглобином.

Особенности переливания крови
В жизни человека часто возникают ситуации, при которых требуется переливание крови. Долгое время такие операции приводили к смерти больных, а настоящие причины этого оставались загадкой. Только в начале 20 века было установлено, что виной всему — эритроцит. Строение этих клеток обусловливает группы крови человека. Всего их четыре, а различают их по системе АВ0.
Каждая из них отличается особым типом белковых веществ, содержащихся в эритроцитах. Называются они агглютиногены. У людей с первой группой крови они отсутствуют. Со второй — имеют агглютиногены А, с третьей — В, с четвертой — АВ. Одновременно в плазме крови содержатся белки агглютинины: альфа, бетта или одновременно оба. Сочетание этих веществ определяет совместимость групп крови. Это значит, что невозможно одновременное присутствие в крови аггглютиногена А и агглютинина альфа. В этом случае эритроциты склеиваются, что может привести к гибели организма.

Что такое резус-фактор
Строение эритроцита человека обусловливает выполнение еще одной функции — определение резус-фактора. Этот признак также обязательно учитывается во время переливания крови. У резус-положительных людей на мембране эритроцита расположен особый белок. Таких людей в мире большинство — более 80 %. У резус — отрицательных людей такого белка нет.
В чем опасность смешивания крови с эритроцитами разных типов? Во время беременности резус-отрицательной женщины в ее кровь могут проникнуть белки плода. В ответ на это организм матери начнет вырабатывать защитные антитела, которые нейтрализуют их. В ходе этого процесса разрушаются эритроциты резус-положительного плода. Современная медицина создала специальные препараты, предотвращающие данный конфликт.
Эритроциты являются красными клетками крови, основной функцией которой является перенос кислорода от легких к клеткам и тканям и углекислого газа в обратном направлении. Выполнение этой роли возможно благодаря двояковогнутой форме, маленьким размерам, высокой концентрации и наличию гемоглобина в клетке.
Костный мозг человека — Википедия
Костный мозг — важнейший орган кроветворной системы, осуществляющий гемопоэз, или кроветворение — процесс создания новых клеток крови взамен погибающих и отмирающих. Он также является одним из органов иммунопоэза. Для иммунной системы человека костный мозг вместе с периферическими лимфоидными органами является функциональным аналогом так называемой фабрициевой сумки, имеющейся у птиц.
Костный мозг — единственная ткань взрослого организма, в норме содержащая большое количество незрелых, недифференцированных и низкодифференцированных клеток, так называемых стволовых клеток, близких по строению к эмбриональным клеткам. Все другие незрелые клетки, например, незрелые клетки кожи, всё же имеют большую степень дифференцировки и зрелости, чем клетки костного мозга, и имеют уже заданную специализацию.
 Клетки красного костного мозга (Gray’s Anatomy)
Клетки красного костного мозга (Gray’s Anatomy)Красный, или кроветворный, костный мозг у человека находится в основном внутри тазовых костей, рёбер, грудины, костей черепа, внутри эпифизов и губчатого вещества эпифизов длинных трубчатых костей и, в ещё меньшей степени, внутри тел позвонков.
Красный костный мозг состоит из фиброзной ткани стромы и собственно кроветворной ткани. В кроветворной ткани костного мозга выделяют несколько ростков гемопоэза (также называемых линиями, англ. cell lines), количество которых увеличивается по мере созревания. Зрелых ростков в красном костном мозге пять: эритроцитарный, гранулоцитарный, лимфоцитарный, моноцитарный и мегакариоцитарный. Каждый из этих ростков даёт, соответственно, следующие клетки и постклеточные элементы: эритроциты; эозинофилы, нейтрофилы и базофилы; лимфоциты; моноциты; тромбоциты.
Развитие ростков гемопоэза представляет собой сложный процесс дифференцировки клеток. Родоначальники всех ростков названы полипотентными клетками за их способность дифференцироваться в клетки всех ростков гемопоэза под действием цитокинов. Также эти клетки называют колониеобразующими элементами (КОЭ) за их локальное расположение в костном мозге. Количество полипотентных стволовых клеток, то есть клеток, которые являются самыми первыми предшественниками в ряду кроветворных клеток, в костном мозге ограничено, и они не могут размножаться, сохраняя полипотентность, и тем самым восстанавливать численность. Дело в том, что при первом же делении полипотентная клетка выбирает путь развития, и её дочерние клетки становятся либо мультипотентными клетками, у которых выбор более ограничен (только в эритроцитарный или лейкоцитарный ростки), либо мегакариобластами и затем мегакариоцитами — клетками, от которых отшнуровываются тромбоциты.

Под действием цитокинов КОЭ начинают специализироваться, переходя на следующий этап — олигопотентные клетки. Как видно из названия, вариантов дифференцировки у них уже меньше. Второе название этих клеток — колониеобразующие единицы (КОЕ), поскольку они расположены более мелкими группами, чем КОЭ. КОЕ неоднородны между собой: выделяют колониеобразующие единицы гранулоцитарно-эритроцитарно-миелоцитарно-макрофагального (КОЕ-ГЭММ) и колониеобразующие единицы лимфоцитарного (КОЕ-Л) ростков. Дальнейшее развитие КОЕ ещё более специфично.
Под действием цитокинов КОЕ-ГЭММ даёт следующие три типа клеток: колониеобразующая единица гранулоцитов и моноцитов (КОЕ-ГиМ), колониеобразующая единица эритроцитов (КОЕ-Э) и колониеобразующая единица мегакариоцитов (КОЕ-МГЦ). Эти переходы инициируются лейкопоэтином, эритропоэтином и тромбопоэтином соответственно. Эти КОЕ — последние, дальнейшие клетки ростков называются бластами, поскольку они уже становятся на один путь дифференцировки в одну конечную клетку. Так, КОЕ-ГМ развивается либо в промонобласт, либо в програнулобласт; КОЕ-Э развивается в эритробласт; КОЕ-МГЦ развивается в мегакариобласт. Таким образом, вкупе с лимфоидным ростком получаются 5 вышеперечисленных ростков гемопоэза.
Под действием различные лимфопоэтинов КОЕ-Л начинает экспрессировать разные маркеры дифференцировки и даёт разные типы лимфоцитов. Например, CD4+, CD8+, CD54+, и так далее. Представления о «Т-лимфоцитах» и «В-лимфоцитах» устарели уже к началу 1990-х годов: это деление основано лишь на месте конечного созревания клетки и не учитывает иммунологических особенностей. Например, классический «Т-лимфоцит» и правда отличается от классического «В-лимфоцита»: первый несёт CD3+, а второй CD19+. Однако такое деление не учитывает множество клеток, не являющихся потомками ни Т-, ни В- ростка. Классическим примером для аргументации неточности этой классификации являются естественные киллеры (NK-клетки). Также такое деление не позволяет объяснять механизмы взаимодействия иммунокомпетентных клеток при различных типах аллергий, что требует искусственно вводить термины «Т-хелперы» и «Т-супрессоры», а в дальнейшем разбивать их на «Т-хелперы 1-го порядка» и «Т-хелперы 2-го порядка». Однако для простоты термины «Т- и В- лимфоциты» используются до сих пор.
Чувствительность к цитостатикам и излучению[править | править код]
Клетки нормального костного мозга, подобно другим незрелым клеткам — клеткам злокачественных опухолей, а также стволовым клеткам кожи и слизистых, — обладают повышенной по сравнению с другими, более зрелыми, клетками организма чувствительностью к ионизирующим излучениям и цитостатическим противоопухолевым химиопрепаратам. Однако чувствительность клеток костного мозга всё же ниже чувствительности клеток злокачественных опухолей, что и позволяет применять химиотерапию и облучение, уничтожая злокачественные опухоли или тормозя их размножение и метастазирование при сравнительно меньшем (хотя во многих случаях и весьма значительном) повреждении костного мозга.
Особенно высокой, более высокой, чем у клеток нормального костного мозга, чувствительностью к химиотерапии обладают лейкозные клетки.
Те цитотоксические химиопрепараты, которые уничтожают или повреждают полипотентные клетки, обладают кумулятивным, то есть накапливающимся, повреждающим воздействием на костномозговое кроветворение. Дело в том, что они приводят к исчерпанию невозобновимого костномозгового резерва первичных клеток-предшественников. Подобный кумулятивный угнетающий эффект на костномозговое кроветворение характерен, в частности, для бусульфана и производных нитрозомочевины. Передозировка любого из цитостатических агентов, обладающих кумулятивным действием на клетки-предшественники, вызывает необратимую аплазию костного мозга — апластическую анемию.
 Проведение пункции.
Проведение пункции.И напротив, химиопрепараты, в основном повреждающие или уничтожающие более поздние промежуточные стадии развития кроветворных клеток, например мультипотентные клетки, почти не обладают кумулятивным угнетающим действием на костномозговое кроветворение — после прекращения химиотерапевтического воздействия численность костномозговых клеточных популяций полностью или почти полностью восстанавливается за счёт костномозгового резерва первичных клеток-предшественников. Таким свойством — относительно мало уничтожать невозобновимую популяцию первичных полипотентных клеток — обладает большинство противоопухолевых препаратов, например, циклофосфамид, цитозин-арабинозид. Именно это позволяет применять эти лекарства при опухолях и лейкозах.
Трансплантация костного мозга
Гемоцитобласт — Википедия
Гемопоэтические стволовые клетки (ГСК, также называемые гемоцитобластами) — это самые ранние предшественники клеток крови, которые дают начало всем остальным клеткам крови и происходят от гемангиобластов и прегемангиобластов, а те, в свою очередь — от клеток первичной эмбриональной мезодермы. Гемопоэтические стволовые клетки находятся в красном костном мозгу, который, в свою очередь, находится внутри полостей большинства костей.
Плюрипотентные гемопоэтические стволовые клетки (гемоцитобласты) дают начало как миелоидным клеткам (моноцитам и макрофагам, трём разновидностям гранулоцитов — нейтрофилам, базофилам и эозинофилам, а также эритроцитам, мегакариоцитам/тромбоцитам, миелоидным дендритным клеткам), так и лимфоидным клеткам (Т-лимфоцитам, B-лимфоцитам, NK-лимфоцитам, лимфоидным дендритным клеткам). Именно в силу такого разнообразия (плюрализма) возможностей для дифференцировки их называют «плюрипотентными» или «универсальными». Ранее считалось, что плюрипотентные гемопоэтические стволовые клетки (гемоцитобласты) — это наиболее ранние из гемопоэтических стволовых клеток, встречающихся в постнатальном (то есть, уже родившемся) и тем более во взрослом организме. Однако в последнее время доказано наличие в постнатальном и даже во взрослом организме небольшого количества ещё более ранних, «пре-гемопоэтических», и ещё более плюрипотентных (то есть имеющих ещё больше различных возможностей для дифференцировки по тому или иному пути) стволовых клеток, могущих стать как гемопоэтическими стволовыми клетками (гемоцитобластами), так и стволовыми клетками эндотелия сосудов (ангиобластами) — так называемых клеток гемогенного эндотелия, гемангиобластов и даже прегемангиобластов. Более того, обнаружилось, что, по-видимому, в некоторых специфических обстоятельствах плюрипотентные гемопоэтические клетки способны к «обратной дифференцировке» — частичной «раздифференцировке» обратно в гемангиобласты и прегемангиобласты (а те, естественно, способны к созреванию в гемоцитобласты). То есть дифференцировка в этой клеточной системе не всегда идёт в одном направлении, от менее дифференцированных клеток к более дифференцированным, и определённое количество гемангиобластов и прегемангиобластов всегда поддерживается во взрослом организме «про запас», при необходимости даже путём «обратной дифференцировки» (которая возможна только на этом этапе, но не на более поздних).
В связи со всеми этими новыми данными определение термина «гемопоэтическая стволовая клетка» сильно изменилось за последние два десятилетия. В настоящее время общепризнано, что гемопоэтические (равно как и пре-гемопоэтические — гемангиобластные) стволовые клетки не представляют собой какую-то единую, однородную гомогенную популяцию одинаковых по своим свойствам клеток, а составляют сложную гетерогенную смесь различных субпопуляций гемопоэтических стволовых клеток разной степени зрелости (находящихся на разных стадиях дифференцировки), и обладающих несколько различными поверхностными антигенами (кластерами дифференцировки), разным временем жизни, разной краткосрочной и долгосрочной регенеративной активностью, разными профилями экспрессии генов и разными эпигенетическими программами дальнейшей дифференцировки, заложенными в них. А гемопоэтическая (кроветворная) ткань костного мозга содержит как собственно ранние гемопоэтические и пре-гемопоэтические стволовые клетки разных субпопуляций и разной степени зрелости, обладающие разными краткосрочными и долгосрочными регенеративными возможностями, так и более поздние, уже полностью или частично коммиттированные (рекрутированные) в ту или иную линию кроветворных клеток, мультипотентные, олигопотентные, бипотентные и унипотентные клетки-предшественники. Собственно, ранние гемопоэтические стволовые клетки (гемоцитобласты) составляют всего лишь 1 из каждых 10 000 клеток в кроветворной (миелоидной) ткани красного костного мозга. Ещё более ранних, пре-гемопоэтических стволовых клеток (гемангиобластов и прегемангиобластов) в кроветворной ткани костного мозга ещё меньше, приблизительно 1: 50 000. Однако чем более ранней является гемопоэтическая стволовая клетка, тем выше её пролиферативная активность и её способность к регенерации, поэтому сравнительно небольшое их количество (в экспериментах на мышах — всего одна гемопоэтическая стволовая клетка) способно реконституировать всю или почти всю кроветворную ткань после её уничтожения сублетальными дозами ионизирующей радиации или цитостатической химиотерапии. На этом основан принцип высокодозной химиотерапии и трансплантации гемопоэтических стволовых клеток.
Как уже было сказано выше, гемопоэтические стволовые клетки являются гетерогенной популяцией, состоящей, на самом деле, из нескольких субпопуляций клеток с разными эпигенетическими программами дальнейшего развития. Так, выделяют три класса гемопоэтических стволовых клеток, различающихся по соотношению лимфоидных и миелоидных клеток (L/M ratio) в популяции их потомков. Миелоидно-уклоняющиеся ГСК (My-bi, от Myeloid-biased) имеют низкое соотношение L/M (0 < L/M < 3), лимфоидно-уклоняющиеся ГСК (Ly-bi, от Lymphoid-biased) имеют высокое соотношение L/M (L/M > 10), а так называемые «сбалансированные» ГСК (Bala, от Balanced) имеют промежуточное соотношение L/M (3 ≤ L/M ≤ 10). Только миелоидно-уклоняющиеся и «сбалансированные» ГСК являются долгоживущими и способны к длительному самовозобновлению популяции. Лимфоидно-уклоняющиеся ГСК — сравнительно короткоживущие. В дополнение к этим данным, эксперименты с последовательной трансплантацией тех или иных типов ГСК показали, что каждый подтип ГСК в условиях другого организма сохраняет свои «предпочтения» в отношении путей дифференцировки и предпочтительным образом воссоздаёт типичное для него распределение типов клеток крови (лимфоидных или миелоидных), характерное для данного подтипа, что заставляет предполагать наличие у этих клеток унаследованной эпигенетической программы дальнейшей дифференцировки для каждого подтипа, на которую оказывает сравнительно малое влияние свойства микроокружения (в частности, внутренняя среда нового организма).
Изучение гемопоэтических стволовых клеток в течение последних 50 лет привело к значительно более глубокому пониманию их свойств, функций и природы. Прогресс в этом понимании сделал возможным широкое применение трансплантации гемопоэтических стволовых клеток при лечении злокачественных опухолей (особенно опухолей системы крови — лейкозов и лимфом), ряда генетических, иммунологических (например, тяжёлый комбинированный иммунодефицит) и гематологических (например, миелодиспластический синдром) заболеваний.[1]
 Схематическое изображение костного мозга и его клеток
Схематическое изображение костного мозга и его клетокГСК обнаруживаются в костном мозге взрослых. Большие их количества находятся в костях таза, бедра и грудины. Также в больших количествах ГСК обнаруживаются в плацентарной и пуповинной крови плода или новорождённого. В небольших количествах ГСК обнаруживаются также в периферической крови здоровых людей.[2] Количество ГСК в периферической крови резко возрастает после введения колониестимулирующих факторов или в фазе восстановления после цитостатической химиотерапии.
Гемопоэтические стволовые клетки и другие клетки-предшественники могут быть взяты из костей таза в области подвздошной кости или из кости грудины, с использованием толстой иглы и шприца. Клетки могут быть изъяты в виде жидкого аспирата, получаемого при отсасывании шприцем (пункционная или аспирационная биопсия костного мозга) или в виде кусочка костномозговой кроветворной ткани, вместе с кусочком стромы костного мозга, костномозговыми сосудами и кусочком кости (так называемая трепанобиопсия костного мозга), при помощи троакара. Трепанобиоптат костного мозга, в отличие от аспирата, позволяет исследовать не только морфологическое строение, иммунофенотип, молекулярную генетику и цитогенетику самих клеток, но и их взаимоотношения друг с другом и с клетками микроокружения (стромы костного мозга, в частности), клетками сосудов и кости, архитектуру костномозговой кроветворной ткани. Это в ряде случаев очень важно для установления диагноза в гематологии.
С целью мобилизации донорских гемопоэтических стволовых клеток из костного мозга в периферическую кровь и их и последующего успешного сбора из периферической крови, донору гемопоэтических стволовых клеток вводят цитокины, такие, как G-CSF и/или GM-CSF, которые, помимо того, что вызывают интенсивное размножение гемопоэтических стволовых клеток в костном мозгу и увеличение их количества, также вызывают их массовый выход из костного мозга в кровь и увеличение количества циркулирующих ГСК.
В эмбриологии млекопитающих, первые явно гемопоэтические стволовые клетки обнаруживаются в области аорта-гонады-мезонефрос. Затем они массивно колонизируют фетальную печень и селезёнку, которые у плода являются основными кроветворными органами. И только затем, сравнительно незадолго до родов, они колонизируют костный мозг и обживаются в нём, и функция органа кроветворения переходит от печени и селезёнки к костному мозгу. Тем не менее, в особых обстоятельствах и у взрослого человека может наблюдаться фетальный (экстрамедуллярный, внекостномозговой) тип кроветворения в селезёнке и в печени. Например, это может случиться при лейкозах, при массивном разрушении костного мозга лейкозными клетками и вытеснении ими из него здоровых гемопоэтических стволовых клеток.[3]
Диаграмма дифференцировки гемопоэтических клеток