Виды РНК, функции и строение
Различные виды ДНК и РНК – нуклеиновых кислот – это один из объектов изучения молекулярной биологии. Одним из наиболее многообещающих и быстро развивающихся направлений в этой науке в последние годы стало исследование РНК.
Кратко о строении РНК
Итак, РНК, рибонуклеиновая кислота, – это биополимер, молекула которого представляет собой цепочку, образованную четырьмя видами нуклеотидов. Каждый нуклеотид, в свою очередь, состоит из азотистого основания (аденина А, гуанина Г, урацила У либо цитозина Ц) в соединении с сахаром рибозой и остатком фосфорной кислоты. Фосфатные остатки, соединяясь с рибозами соседних нуклеотидов, «сшивают» составные блоки РНК в макромолекулу – полинуклеотид. Так образуется первичная структура РНК.

Вторичная структура – образование двойной цепочки – образуется на некоторых участках молекулы в соответствии с принципом комплементарности азотистых оснований: аденин образует пару с урацилом посредством двойной, а гуанин с цитозином — тройной водородной связи.
В рабочей форме молекула РНК образует также третичную структуру – особое пространственное строение, конформацию.
Синтез РНК
Все виды РНК синтезируются при помощи фермента РНК-полимеразы. Она может быть ДНК- и РНК-зависимой, то есть катализировать синтез как на ДНК, так и на РНК-матрице.
Синтез основан на комплементарности оснований и антипараллельности направления чтения генетического кода и протекает в несколько этапов.
Сначала происходит узнавание и связывание РНК-полимеразы с особой последовательностью нуклеотидов на ДНК – промотором, после чего двойная спираль ДНК раскручивается на небольшом участке и начинается сборка молекулы РНК над одной из цепочек, называемой матричной (другая цепочка ДНК называется кодирующей – именно ее копией является синтезируемая РНК). Асимметричность промотора определяет, какая из цепочек ДНК будет служить матрицей, и тем самым позволяет РНК-полимеразе инициировать синтез в правильном направлении.
Следующий этап называется элонгацией. Транскрипционный комплекс, включающий РНК-полимеразу и расплетенный участок с гибридом ДНК-РНК, начинает движение. По мере этого перемещения наращиваемая цепочка РНК постепенно отделяется, а двойная спираль ДНК расплетается перед комплексом и восстанавливается за ним.

Завершающий этап синтеза наступает, когда РНК-полимераза достигает особого участка матрицы, называемого терминатором. Терминация (окончание) процесса может достигаться различными способами.
Основные виды РНК и их функции в клетке
Они следующие:
- Матричная или информационная (мРНК). Посредством ее осуществляется транскрипция – перенос генетической информации с ДНК.
- Рибосомная (рРНК), обеспечивающая процесс трансляции – синтез белка на матрице мРНК.
- Транспортная (тРНК). Производит узнавание и транспортировку аминокислоты на рибосому, где происходит синтез белка, а также принимает участие в трансляции.
- Малые РНК – обширный класс молекул небольшой длины, осуществляющих разнообразные функции в ходе процессов транскрипции, созревания РНК, трансляции.
- РНК-геномы – кодирующие последовательности, которые содержат генетическую информацию у некоторых вирусов и вироидов.
В 1980-х годах была открыта каталитическая активность РНК. Молекулы, обладающие этим свойством, получили название рибозимов. Естественных рибозимов пока известно не так много, каталитическая способность их ниже, чем у белков, однако в клетке они выполняют исключительно важные функции. В настоящее время ведутся успешные работы по синтезу рибозимов, имеющие в том числе и прикладное значение.
Остановимся подробнее на различных видах молекул РНК.
Матричная (информационная) РНК
Эта молекула синтезируется над расплетенным участком ДНК, копируя таким образом ген, кодирующий тот или иной белок.
РНК эукариотических клеток, прежде чем стать, в свою очередь, матрицей для синтеза белка, должны созреть, то есть пройти через комплекс различных модификаций – процессинг.
Прежде всего, еще на стадии транскрипции, молекула подвергается кэпированию: к ее концу присоединяется особая структура из одного или нескольких модифицированных нуклеотидов – кэп. Он играет важную роль во многих последующих процессах и повышает стабильность мРНК. К другому концу первичного транскрипта присоединяется так называемый поли(А)хвост – последовательность адениновых нуклеотидов.
После этого пре-мРНК подвергается сплайсингу. Это удаление из молекулы некодирующих участков – интронов, которых много в ДНК эукариот. Далее происходит процедура редактирования мРНК, при которой химически модифицируется ее состав, а также метилирование, после чего зрелая мРНК покидает клеточное ядро.

Рибосомная РНК
Основу рибосомы – комплекса, обеспечивающего белковый синтез, составляют две длинные рРНК, которые образуют субчастицы рибосомы. Синтезируются они совместно в виде одной пре-рРНК, которая затем в ходе процессинга разделяется. В большую субчастицу входит также низкомолекулярная рРНК, синтезируемая с отдельного гена. Рибосомные РНК обладают плотно упакованной третичной структурой, которая служит каркасом для белков, присутствующих в рибосоме и выполняющих вспомогательные функции.
В нерабочей фазе субъединицы рибосомы разделены; при инициации трансляционного процесса рРНК малой субчастицы соединяется с матричной РНК, после чего происходит полное объединение элементов рибосомы. При взаимодействии РНК малой субчастицы с мРНК последняя как бы протягивается через рибосому (что равнозначно движению рибосомы по мРНК). Рибосомная РНК большой субчастицы является рибозимом, то есть обладает ферментными свойствами. Она катализирует образование пептидных связей между аминокислотами в ходе синтеза белка.

Следует отметить, что наибольшая часть всей РНК в клетке приходится на долю рибосомной – 70-80 %. ДНК обладает большим количеством генов, кодирующих рРНК, что обеспечивает весьма интенсивную ее транскрипцию.
Транспортная РНК
Эта молекула распознается определенной аминокислотой при помощи особого фермента и, соединяясь с ней, производит транспортировку аминокислоты на рибосому, где служит посредником в процессе трансляции – синтеза белка. Перенос осуществляется путем диффузии в цитоплазме клетки.
Вновь синтезированные молекулы тРНК, так же как и другие виды РНК, подвергаются процессингу. Зрелая тРНК в активной форме имеет конформацию, напоминающую клеверный лист. На «черешке» листа – акцепторном участке – расположена последовательность ЦЦА с гидроксильной группой, которая связывается с аминокислотой. На противоположном конце «листа» находится антикодоновая петля, которая соединяется с комплементарным кодоном на мРНК. D-петля служит для связывания транспортной РНК с ферментом при взаимодействии с аминокислотой, а Т-петля – для связывания с большой субчастицей рибосомы.

Малые РНК
Эти виды РНК играют важную роль в клеточных процессах и сейчас активно изучаются.
Так, например, малые ядерные РНК в клетках эукариот участвуют в сплайсинге мРНК и, возможно, обладают каталитическими свойствами наряду с белками сплайсосом. Малые ядрышковые РНК участвуют в процессинге рибосомной и транспортной РНК.
Малые интерферирующие и микроРНК являются важнейшими элементами системы регуляции экспрессии генов, необходимой клетке для контроля собственной структуры и жизнедеятельности. Эта система – важная часть иммунного антивирусного ответа клетки.
Существует также класс малых РНК, функционирующих в комплексе с белками Piwi. Эти комплексы играют огромную роль в развитии клеток зародышевой линии, в сперматогенезе и в подавлении мобильных генетических элементов.
РНК-геном
Молекула РНК может использоваться в качестве генома большинством вирусов. Вирусные геномы бывают различными – одно- и двухцепочечными, кольцевыми или линейными. Также РНК-геномы вирусов часто бывают сегментированы и в целом короче, чем ДНК-содержащие геномы.
Существует семейство вирусов, генетическая информация которых, закодированная в РНК, после инфицирования клетки путем обратной транскрипции переписывается на ДНК, которая затем внедряется в геном клетки-жертвы. Это так называемые ретровирусы. К ним, в частности, относится вирус иммунодефицита человека.

Значение исследования РНК в современной науке
Если прежде преобладало мнение о второстепенной роли РНК, то ныне ясно, что она — необходимый и важнейший элемент внутриклеточной жизнедеятельности. Множество процессов первостепенной значимости не обходятся без активного участия РНК. Механизмы таких процессов долгое время оставались неизвестными, но благодаря исследованию различных видов РНК и их функций постепенно проясняются многие детали.
Не исключено, что РНК сыграла решающую роль в возникновении и становлении жизни на заре истории Земли. Результаты недавних исследований говорят в пользу этой гипотезы, свидетельствуя о необычайной древности многих механизмов функционирования клетки с участием тех или иных видов РНК. Например, недавно открытые рибопереключатели в составе мРНК (система безбелковой регуляции активности генов на стадии транскрипции), по мнению многих исследователей, являются отголосками эпохи, когда примитивная жизнь строилась на основе РНК, без участия ДНК и белков. Также весьма древним компонентом системы регуляции считаются микроРНК. Особенности структуры каталитически активной рРНК свидетельствуют о ее постепенной эволюции путем присоединения новых фрагментов к древней проторибосоме.
Тщательное изучение того, какие виды РНК и каким образом заняты в тех или иных процессах, исключительно важно также для теоретических и прикладных областей медицины.
Дезоксирибонуклеиновая кислота — Википедия
Структура ДНК (двойная спираль). Различные атомы в структуре показаны в разных цветах; детальная структура двух пар оснований показана снизу справа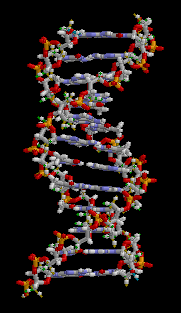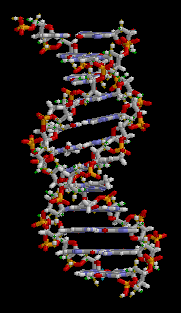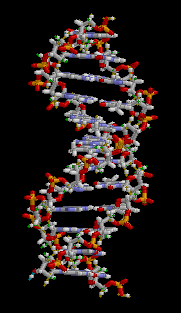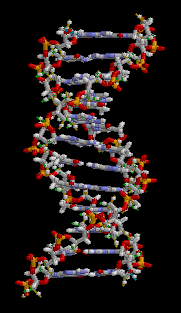
Дезоксирибонуклеи́новая кислота́ (ДНК) — макромолекула (одна из трёх основных, две другие — РНК и белки), обеспечивающая хранение, передачу из поколения в поколение и реализацию генетической программы развития и функционирования живых организмов. Молекула ДНК хранит биологическую информацию в виде генетического кода, состоящего из последовательности нуклеотидов [1]. ДНК содержит информацию о структуре различных видов РНК и белков.
В клетках эукариот (животных, растений и грибов) ДНК находится в ядре клетки в составе хромосом, а также в некоторых клеточных органеллах (митохондриях и пластидах). В клетках прокариотических организмов (бактерий и архей) кольцевая или линейная молекула ДНК, так называемый нуклеоид, прикреплена изнутри к клеточной мембране. У них и у низших эукариот (например дрожжей) встречаются также небольшие автономные, преимущественно кольцевые молекулы ДНК, называемые плазмидами. Кроме того, одно- или двухцепочечные молекулы ДНК могут образовывать геном ДНК-содержащих вирусов.
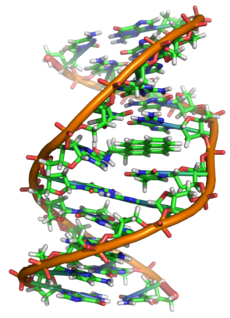
С химической точки зрения ДНК — длинная полимерная молекула, состоящая из повторяющихся блоков — нуклеотидов. Каждый нуклеотид состоит из азотистого основания, сахара (дезоксирибозы) и фосфатной группы. Связи между нуклеотидами в цепи образуются за счёт дезоксирибозы и фосфатной группы (фосфодиэфирные связи). В подавляющем большинстве случаев (кроме некоторых вирусов, содержащих одноцепочечную ДНК) макромолекула ДНК состоит из двух цепей, ориентированных азотистыми основаниями друг к другу. Эта двухцепочечная молекула закручена по винтовой линии. В целом структура молекулы ДНК получила традиционное, но ошибочное название «двойной спирали», на самом же деле она является «двойным винтом». Винтовая линия может быть правой (A- и B-формы ДНК) или левой (Z-форма ДНК)
В ДНК встречается четыре вида азотистых оснований (аденин (A), гуанин (G), тимин (T) и цитозин (C)). Азотистые основания одной из цепей соединены с азотистыми основаниями другой цепи водородными связями согласно принципу комплементарности: аденин (A) соединяется только с тимином (T), гуанин (G) — только с цитозином (C). Последовательность нуклеотидов позволяет «кодировать» информацию о различных типах РНК, наиболее важными из которых являются информационные, или матричные (мРНК), рибосомальные (рРНК) и транспортные (тРНК). Все эти типы РНК синтезируются на матрице ДНК за счёт копирования последовательности ДНК в последовательность РНК, синтезируемой в процессе транскрипции, и принимают участие в биосинтезе белков (процессе трансляции). Помимо кодирующих последовательностей, ДНК клеток содержит последовательности, выполняющие регуляторные и структурные функции. Кроме того, в геноме эукариот часто встречаются участки, принадлежащие «генетическим паразитам», например транспозонам.
Расшифровка структуры ДНК (1953 год) стала одним из поворотных моментов в истории биологии. За выдающийся вклад в это открытие Фрэнсису Крику, Джеймсу Уотсону и Морису Уилкинсу была присуждена Нобелевская премия по физиологии или медицине 1962 года. Розалинд Франклин, которая получила рентгенограммы, без которых Уотсон и Крик не имели бы возможность сделать выводы о структуре ДНК, умерла в 1958 году от рака (Нобелевскую премию не дают посмертно)




ДНК как химическое вещество была выделена Иоганном Фридрихом Мишером в 1869 году из остатков клеток, содержащихся в гное. Он выделил вещество, в состав которого входят азот и фосфор. Вначале новое вещество получило название нуклеин, а позже, когда Мишер определил, что это вещество обладает кислотными свойствами, вещество получило название нуклеиновая кислота[4]. Биологическая функция новооткрытого вещества была неясна, и долгое время ДНК считалась запасником фосфора в организме. Более того, даже в начале XX века многие биологи считали, что ДНК не имеет никакого отношения к передаче информации, поскольку строение молекулы, по их мнению, было слишком однообразным и не могло содержать закодированную информацию.
До 1930-х годов считалось, что ДНК содержится только в животных клетках, а в растительных — РНК. В 1934 году в журнале «Hoppe-Seyler’s Zeitschrift fur physiologishe Chemie»[5], затем в 1935 году в «Ученых записках МГУ»[6] вышла статья советских биохимиков А. Н. Белозерского и А. Р. Кизеля в которых доказывалось присутствие ДНК в растительных клетках. В 1936 году группой Белозерского ДНК была выделена из семян и тканей бобовых, злаковых и других растений[7]. Результатом исследований этой же группы советских учёных в 1939 – 1947 годах стала первая в мировой научной литературе информация о содержании нуклеиновых кислот у различных видов бактерий.
Постепенно было доказано, что именно ДНК, а не белки, как считалось раньше, является носителем генетической информации. Одно из первых решающих доказательств принесли эксперименты Освальда Эвери, Колина Маклауда и Маклина Маккарти (1944 г.) по трансформации бактерий. Им удалось показать, что за так называемую трансформацию (приобретение болезнетворных свойств безвредной культурой в результате добавления в неё мёртвых болезнетворных бактерий) отвечает выделенная из пневмококков ДНК. Эксперимент американских учёных Алфреда Херши и Марты Чейз (эксперимент Херши — Чейз, 1952 г.) с помеченными радиоактивными изотопами белками и ДНК бактериофагов показали, что в заражённую клетку передаётся только нуклеиновая кислота фага, а новое поколение фага содержит такие же белки и нуклеиновую кислоту, как исходный фаг[8]
.Вплоть до 50-х годов XX века точное строение ДНК, как и способ передачи наследственной информации, оставалось неизвестным. Хотя и было доподлинно известно, что ДНК состоит из нескольких цепочек, состоящих из нуклеотидов, никто не знал точно, сколько этих цепочек и как они соединены.
В результате работы группы биохимика Эрвина Чаргаффа в 1949—1951 гг. были сформулированы так называемые правила Чаргаффа. Чаргаффу и сотрудникам удалось разделить нуклеотиды ДНК при помощи бумажной хроматографии и определить точные количественные соотношения нуклеотидов разных типов. Соотношение, выявленное для аденина (А), тимина (Т), гуанина (Г) и цитозина (Ц), оказалось следующим: количество аденина равно количеству тимина, а гуанина — цитозину: А=Т, Г=Ц[9][10]. Эти правила, наряду с данными рентгеноструктурного анализа, сыграли решающую роль в расшифровке структуры ДНК.
Структура двойной спирали ДНК была предложена Френсисом Криком и Джеймсом Уотсоном в 1953 году на основании рентгеноструктурных данных, полученных Морисом Уилкинсом и Розалинд Франклин, и правил Чаргаффа
Интересно, что в 1957 году американцы Александер Рич, Гэри Фелзенфелд и Дэйвид Дэйвис описали нуклеиновую кислоту, составленную тремя спиралями[13]. А в 1985—1986 годах Максим Давидович Франк-Каменецкий в Москве показал, как двухспиральная ДНК складывается в так называемую H-форму, составленную уже не двумя, а тремя нитями ДНК[14][15].
Нуклеотиды[править | править код]
Структуры оснований в составе ДНК
Дезоксирибонуклеиновая кислота (ДНК) представляет собой биополимер (полианион), мономером которого является нуклеотид[16][17]
.Каждый нуклеотид состоит из остатка фосфорной кислоты, присоединённого по 5′-положению к сахару дезоксирибозе, к которому также через гликозидную связь (C—N) по 1′-положению присоединено одно из четырёх азотистых оснований. Именно наличие характерного сахара и составляет одно из главных различий между ДНК и РНК, зафиксированное в названиях этих нуклеиновых кислот (в состав РНК входит сахар рибоза)[18]. Пример нуклеотида — аденозинмонофосфат, у которого основанием, присоединённым к фосфату и рибозе, является аденин (A) (показан на рисунке).
Исходя из структуры молекул, основания, входящие в состав нуклеотидов, разделяют на две группы: пурины (аденин [A] и гуанин [G]) образованы соединёнными пяти- и шестичленным гетероциклами; пиримидины (цитозин [C] и тимин [T]) — шестичленным гетероциклом[19].
В виде исключения, например, у бактериофага PBS1, в ДНК встречается пятый тип оснований — урацил ([U]), пиримидиновое основание, отличающееся от тимина отсутствием метильной группы на кольце, обычно заменяющее тимин в РНК
Следует отметить, что тимин (T) и урацил (U) не так строго приурочены к ДНК и РНК соответственно, как это считалось ранее. Так, после синтеза некоторых молекул РНК значительное число урацилов в этих молекулах метилируется с помощью специальных ферментов, превращаясь в тимин. Это происходит в транспортных и рибосомальных РНК[21].
Двойная спираль[править | править код]
В зависимости от концентрации ионов и нуклеотидного состава молекулы, двойная спираль ДНК в живых организмах существует в разных формах. На рисунке представлены формы A, B и Z (слева направо)Полимер ДНК обладает довольно сложной структурой. Нуклеотиды соединены между собой ковалентно в длинные полинуклеотидные цепи. Эти цепи в подавляющем большинстве случаев (кроме некоторых вирусов, обладающих одноцепочечными ДНК-геномами) попарно объединяются при помощи водородных связей во вторичную структуру, получившую название двойной спирали[11][18]. Остов каждой из цепей состоит из чередующихся фосфатов и сахаров[22]. Внутри одной цепи ДНК соседние нуклеотиды соединены фосфодиэфирными связями, которые формируются в результате взаимодействия между 3′-гидроксильной (3’—ОН) группой молекулы дезоксирибозы одного нуклеотида и 5′-фосфатной группой (5’—РО3) другого. Асимметричные концы цепи ДНК называются 3′ (три прайм) и 5′ (пять прайм). Полярность цепи играет важную роль при синтезе ДНК (удлинение цепи возможно только путём присоединения новых нуклеотидов к свободному 3′-концу).
Как уже было сказано выше, у подавляющего большинства живых организмов ДНК состоит не из одной, а из двух полинуклеотидных цепей. Эти две длинные цепи закручены одна вокруг другой в виде двойной спирали, стабилизированной водородными связями, образующимися между обращёнными друг к другу азотистыми основаниями входящих в неё цепей. В природе эта спираль, чаще всего, правозакрученная. Направления от 3′-конца к 5′-концу в двух цепях, из которых состоит молекула ДНК, противоположны (цепи «антипараллельны» друг другу).
Ширина двойной спирали составляет от 22 до 24 Å, или 2,2—2,4 нм, длина каждого нуклеотида 3,3 Å (0,33 нм)[23]. Подобно тому, как в винтовой лестнице сбоку можно увидеть ступеньки, на двойной спирали ДНК в промежутках между фосфатным остовом молекулы можно видеть рёбра оснований, кольца которых расположены в плоскости, перпендикулярной по отношению к продольной оси макромолекулы.
В двойной спирали различают малую (12 Å) и большую (22 Å) бороздки[24]. Белки, например, факторы транскрипции, которые присоединяются к определённым последовательностям в двухцепочечной ДНК, обычно взаимодействуют с краями оснований в большой бороздке, где те более доступны[25].
Образование связей между основаниями[править | править код]
Каждое основание на одной из цепей связывается с одним определённым основанием на второй цепи. Такое специфическое связывание называется комплементарным. Пурины комплементарны пиримидинам (то есть способны к образованию водородных связей с ними): аденин образует связи только с тимином, а цитозин — с гуанином. В двойной спирали цепочки также связаны с помощью гидрофобных взаимодействий и стэкинга, которые не зависят от последовательности оснований ДНК[26].
Комплементарность двойной спирали означает, что информация, содержащаяся в одной цепи, содержится и в другой цепи. Обратимость и специфичность взаимодействий между комплементарными парами оснований важна для репликации ДНК и всех остальных функций ДНК в живых организмах.
Так как водородные связи нековалентны, они легко разрываются и восстанавливаются. Цепочки двойной спирали могут расходиться как замок-молния под действием ферментов (хеликазы) или при высокой температуре[27]. Разные пары оснований образуют разное количество водородных связей. АТ связаны двумя, ГЦ — тремя водородными связями, поэтому на разрыв ГЦ требуется больше энергии. Процент ГЦ-пар и длина молекулы ДНК определяют количество энергии, необходимой для диссоциации цепей: длинные молекулы ДНК с большим содержанием ГЦ более тугоплавки[28].
Части молекул ДНК, которые из-за их функций должны быть легко разделяемы, например, ТАТА последовательность в бактериальных промоторах, обычно содержат большое количество А и Т.
Химические модификации азотистых оснований[править | править код]
Структура цитозина, 5-метилцитозина и тимина. Тимин может возникать путём деаминирования 5-метилцитозина
Азотистые основания в составе ДНК могут быть ковалентно модифицированы, что используется при регуляции экспрессии генов. Например, в клетках позвоночных метилирование цитозина с образованием 5-метилцитозина используется соматическими клетками для передачи профиля генной экспрессии дочерним клеткам. Метилирование цитозина не влияет на спаривание оснований в двойной спирали ДНК. У позвоночных метилирование ДНК в соматических клетках ограничивается метилированием цитозина в последовательности ЦГ[29]. Средний уровень метилирования отличается у разных организмов, так, у нематоды Caenorhabditis elegans метилирование цитозина не наблюдается, а у позвоночных обнаружен высокий уровень метилирования — до 1 %[30]. Другие модификации оснований включают метилирование аденина у бактерий и гликозилирование урацила с образованием «J-основания» в кинетопластах[31].
Метилирование цитозина с образованием 5-метилцитозина в промоторной части гена коррелирует с его неактивным состоянием[32]. Метилирование цитозина важно также для инактивации Х-хромосомы у млекопитающих[33]. Метилирование ДНК используется в геномном импринтинге[34]. Значительные нарушения профиля метилирования ДНК происходят при канцерогенезе[35].
Несмотря на биологическую роль, 5-метилцитозин может спонтанно утрачивать аминную группу (деаминироваться), превращаясь в тимин, поэтому метилированные цитозины являются источником повышенного числа мутаций[36].
Повреждения ДНК[править | править код]
 Интеркалированное химическое соединение, которое находится в середине спирали — бензопирен, основной мутаген табачного дыма[37]
Интеркалированное химическое соединение, которое находится в середине спирали — бензопирен, основной мутаген табачного дыма[37]ДНК может повреждаться разнообразными мутагенами, к которым относятся окисляющие и алкилирующие вещества, а также высокоэнергетическая электромагнитная радиация — ультрафиолетовое и рентгеновское излучение. Тип повреждения ДНК зависит от типа мутагена. Например, ультрафиолет повреждает ДНК путём образования в ней димеров тимина, которые возникают при образовании ковалентных связей между соседними основаниями[38].
Оксиданты, такие как свободные радикалы или пероксид водорода, приводят к нескольким типам повреждения ДНК, включая модификации оснований, в особенности гуанозина, а также двухцепочечные разрывы в ДНК[39]. По некоторым оценкам, в каждой клетке человека окисляющими соединениями ежедневно повреждается порядка 500 оснований[40][41]. Среди разных типов повреждений наиболее опасные — это двухцепочечные разрывы, потому что они трудно репарируются и могут привести к потерям участков хромосом (делециям) и транслокациям.
Многие молекулы мутагенов вставляются (интеркалируют) между двумя соседними парами оснований. Большинство этих соединений, например: бромистый этидий, даунорубицин, доксорубицин и талидомид, имеет ароматическую структуру. Для того чтобы интеркалирующее соединение могло поместиться между основаниями, они должны разойтись, расплетая и нарушая структуру двойной спирали. Эти изменения в структуре ДНК мешают транскрипции и репликации, вызывая мутации. Поэтому интеркалирующие соединения часто являются канцерогенами, наиболее известные из которых — бензопирен, акридины, афлатоксин и бромистый этидий[42][43][44]. Несмотря на эти негативные свойства, в силу их способности подавлять транскрипцию и репликацию ДНК, интеркалирующие соединения используются в химиотерапии для подавления быстро растущих клеток рака[45].
Некоторые вещества (цисплатин[46], митомицин C[47], псорален[48]) образуют поперечные сшивки между нитями ДНК и подавляют синтез ДНК, благодаря чему используются в химиотерапии некоторых видов рака (см. Химиотерапия злокачественных новообразований).
Суперскрученность[править | править код]
Если взяться за концы верёвки и начать скручивать их в разные стороны, она становится короче и на верёвке образуются «супервитки». Так же может быть суперскручена и ДНК. В обычном состоянии цепочка ДНК делает один оборот на каждые 10,459 основания, но в суперскрученном состоянии спираль может быть свёрнута туже или расплетена[49]. Выделяют два типа суперскручивания: положительное — в направлении нормальных витков, при котором основания расположены ближе друг к другу; и отрицательное — в противоположном направлении. В природе молекулы ДНК обычно находятся в отрицательном суперскручивании, которое вносится ферментами — топоизомеразами[50]. Эти ферменты удаляют дополнительное скручивание, возникающее в ДНК в результате транскрипции и репликации[51].
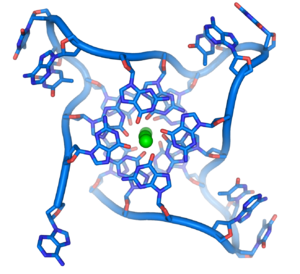 Структура теломер. Зелёным цветом показан ион металла, хелатированный в центре структуры[52]
Структура теломер. Зелёным цветом показан ион металла, хелатированный в центре структуры[52]Структуры на концах хромосом[править | править код]
На концах линейных хромосом находятся специализированные структуры ДНК, называемые теломерами. Основная функция этих участков — поддержание целостности концов хромосом[53]. Теломеры также защищают концы ДНК от деградации экзонуклеазами и предотвращают активацию системы репарации[54]. Поскольку обычные ДНК-полимеразы не могут реплицировать 3′ концы хромосом, это делает специальный фермент — теломераза.
В клетках человека теломеры часто представлены одноцепочечной ДНК и состоят из нескольких тысяч повторяющихся единиц последовательности ТТАГГГ[55]. Эти последовательности с высоким содержанием гуанина стабилизируют концы хромосом, формируя очень необычные структуры, называемые G-квадруплексами и состоящие из четырёх, а не двух взаимодействующих оснований. Четыре гуаниновых основания, все атомы которых находятся в одной плоскости, образуют пластинку, стабилизированную водородными связями между основаниями и хелатированием в центре неё иона металла (чаще всего калия). Эти пластинки располагаются стопкой друг над другом[56].
На концах хромосом могут образовываться и другие структуры: основания могут быть расположены в одной цепочке или в разных параллельных цепочках. Кроме этих «стопочных» структур теломеры формируют большие петлеобразные структуры, называемые Т-петли или теломерные петли. В них одноцепочечная ДНК располагается в виде широкого кольца, стабилизированного теломерными белками[57]. В конце Т-петли одноцепочечная теломерная ДНК присоединяется к двухцепочечной ДНК, нарушая спаривание цепочек в этой молекуле и образуя связи с одной из цепей. Это трёхцепочечное образование называется Д-петля (от англ. displacement loop)[56].
ДНК является носителем генетической информации, записанной в виде последовательности нуклеотидов с помощью генетического кода. С молекулами ДНК связаны два основополагающих свойства живых организмов — наследственность и изменчивость. В ходе процесса, называемого репликацией ДНК, образуются две копии исходной цепочки, наследуемые дочерними клетками при делении, отсюда следует, что образовавшиеся клетки оказываются генетически идентичны исходной.
Генетическая информация реализуется при экспрессии генов в процессах транскрипции (синтеза молекул РНК на матрице ДНК) и трансляции (синтеза белков на матрице РНК).
Последовательность нуклеотидов «кодирует» информацию о различных типах РНК: информационных, или матричных (мРНК), рибосомальных (рРНК) и транспортных (тРНК). Все эти типы РНК синтезируются на основе ДНК в процессе транскрипции. Роль их в биосинтезе белков (процессе трансляции) различна. Информационная РНК содержит информацию о последовательности аминокислот в белке, рибосомальные РНК служат основой для рибосом (сложных нуклеопротеиновых комплексов, основная функция которых — сборка белка из отдельных аминокислот на основе иРНК), транспортные РНК доставляют аминокислоты к месту сборки белков — в активный центр рибосомы, «ползущей» по иРНК.
Структура генома[править | править код]
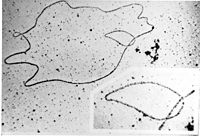 ДНК генома бактериофага: фотография под просвечивающим электронным микроскопом
ДНК генома бактериофага: фотография под просвечивающим электронным микроскопомБольшинство природных ДНК имеет двухцепочечную структуру, линейную (эукариоты, некоторые вирусы и отдельные роды бактерий) или кольцевую (прокариоты, хлоропласты и митохондрии). Линейную одноцепочечную ДНК содержат некоторые вирусы и бактериофаги. Молекулы ДНК находятся in vivo в плотно упакованном, конденсированном состоянии[58]. В клетках эукариот ДНК располагается главным образом в ядре и на стадии профазы, метафазы или анафазы митоза доступны для наблюдения с помощью светового микроскопа в виде набора хромосом. Бактериальная (прокариоты) ДНК обычно представлена одной кольцевой молекулой ДНК, расположенной в неправильной формы образовании в цитоплазме, называемым нуклеоидом[59]. Генетическая информация генома состоит из генов. Ген — единица передачи наследственной информации и участок ДНК, который влияет на определённую характеристику организма. Ген содержит открытую рамку считывания, которая транскрибируется, а также регуляторные последовательности (англ.)русск., например промотор и энхансер, которые контролируют экспрессию открытых рамок считывания.
У многих видов только малая часть общей последовательности генома кодирует белки. Так, только около 1,5 % генома человека состоит из кодирующих белок экзонов, а больше 50 % ДНК человека состоит из некодирующих повторяющихся последовательностей ДНК[60]. Причины наличия такого большого количества некодирующей ДНК в эукариотических геномах и огромная разница в размерах геномов (С-значение) — одна из неразрешённых научных загадок[61]; исследования в этой области также указывают на большое количество фрагментов реликтовых вирусов в этой части ДНК.
Последовательности генома, не кодирующие белок[править | править код]
В настоящее время накапливается всё больше данных, противоречащих идее о некодирующих последовательностях как «мусорной ДНК» (англ. junk DNA). Теломеры и центромеры содержат малое число генов, но они важны для функционирования и стабильности хромосом[54][62]. Часто встречающаяся форма некодирующих последовательностей человека — псевдогены, копии генов, инактивированные в результате мутаций[63]. Эти последовательности нечто вроде молекулярных ископаемых, хотя иногда они могут служить исходным материалом для дупликации и последующей дивергенции генов[64]. Другой источник разнообразия белков в организме — это использование интронов в качестве «линий разреза и склеивания» в альтернативном сплайсинге[65]. Наконец, не кодирующие белок последовательности могут кодировать вспомогательные клеточные РНК, например мяРНК[66]. Недавнее исследование транскрипции генома человека показало, что 10 % генома даёт начало полиаденилированным РНК[67], а исследование генома мыши показало, что 62 % его транскрибируется[68].
Транскрипция и трансляция[править | править код]
Генетическая информация, закодированная в ДНК, должна быть прочитана и в конечном итоге выражена в синтезе различных биополимеров, из которых состоят клетки. Последовательность оснований в цепочке ДНК напрямую определяет последовательность оснований в РНК, на которую она «переписывается» в процессе, называемом транскрипцией. В случае мРНК эта последовательность определяет аминокислоты белка. Соотношение между нуклеотидной последовательностью мРНК и аминокислотной последовательностью определяется правилами трансляции, которые называются генетическим кодом. Генетический код состоит из трёхбуквенных «слов», называемых кодонами, состоящих из трёх нуклеотидов (то есть ACT, CAG, TTT и т. п.). Во время транскрипции нуклеотиды гена копируются на синтезируемую РНК РНК-полимеразой. Эта копия в случае мРНК декодируется рибосомой, которая «читает» последовательность мРНК, осуществляя спаривание матричной РНК с транспортными РНК, которые присоединены к аминокислотам. Поскольку в трёхбуквенных комбинациях используются 4 основания, всего возможны 64 кодона (4³ комбинации). Кодоны кодируют 20 стандартных аминокислот, каждой из которых соответствует в большинстве случаев более одного кодона. Один из трёх кодонов, которые располагаются в конце мРНК, не означает аминокислоту и определяет конец белка, это «стоп» или «нонсенс» кодоны — TAA, TGA, TAG.
Репликация[править | править код]
Деление клеток необходимо для размножения одноклеточного и роста многоклеточного организма, но до деления клетка должна удвоить геном, чтобы дочерние клетки содержали ту же генетическую информацию, что и исходная клетка. Из нескольких теоретически возможных механизмов удвоения (репликации) ДНК реализуется полуконсервативный. Две цепочки разделяются, а затем каждая недостающая комплементарная последовательность ДНК воспроизводится ферментом ДНК-полимеразой. Этот фермент синтезирует полинуклеотидную цепь, находя правильный нуклеотид через комплементарное спаривание оснований и присоединяя его к растущей цепочке. ДНК-полимераза не может начинать новую цепь, а может лишь наращивать уже существующую, поэтому она нуждается в короткой цепочке нуклеотидов — (праймере), синтезируемом праймазой. Так как ДНК-полимеразы могут синтезировать цепочку только в направлении 5′ —> 3′, антипараллельные цепи ДНК копируются по-разному: одна цепь синтезируется непрерывно, а вторая прерывчато[69].
Все функции ДНК зависят от её взаимодействия с белками. Взаимодействия могут быть неспецифическими, когда белок присоединяется к любой молекуле ДНК, или зависеть от наличия особой последовательности. Ферменты также могут взаимодействовать с ДНК, из них наиболее важные — это РНК-полимеразы, которые копируют последовательность оснований ДНК на РНК в транскрипции или при синтезе новой цепи ДНК — репликации.
Структурные и регуляторные белки[править | править код]
Хорошо изученными примерами взаимодействия белков и ДНК, не зависящего от нуклеотидной последовательности ДНК, является взаимодействие со структурными белками. В клетке ДНК связана с этими белками, образуя компактную структуру, которая называется хроматин. У эукариот хроматин образован при присоединении к ДНК небольших щелочных белков — гистонов, менее упорядоченный хроматин прокариот содержит гистон-подобные белки[70][71]. Гистоны формируют дискообразную белковую структуру — нуклеосому, вокруг каждой из которых вмещается два оборота спирали ДНК. Неспецифические связи между гистонами и ДНК образуются за счёт ионных связей щелочных аминокислот гистонов и кислотных остатков сахарофосфатного остова ДНК[72]. Химические модификации этих аминокислот включают метилирование, фосфорилирование и ацетилирование[73]. Эти химические модификации изменяют силу взаимодействия между ДНК и гистонами, влияя на доступность специфических последовательностей для факторов транскрипции и изменяя скорость транскрипции[74]. Другие белки в составе хроматина, которые присоединяются к неспецифическим последовательностям — белки с высокой подвижностью в гелях, которые ассоциируют большей частью с согнутой ДНК[75]. Эти белки важны для образования в хроматине структур более высокого порядка[76].
Особая группа белков, присоединяющихся к ДНК — это белки, которые ассоциируют с одноцепочечной ДНК. Наиболее хорошо охарактеризованный белок этой группы у человека — репликационный белок А, без которого невозможно протекание большинства процессов, где расплетается двойная спираль, включая репликацию, рекомбинацию и репарацию. Белки этой группы стабилизируют одноцепочечную ДНК и предотвращают формирование стеблей-петель или деградации нуклеазами[77].
В то же время другие белки узнают и присоединяются к специфическим последовательностям. Наиболее изученная группа таких белков — различные классы факторов транскрипции, то есть белки, регулирующие транскрипцию. Каждый из этих белков узнаёт свою последовательность, часто в промоторе, и активирует или подавляет транскрипцию гена. Это происходит при ассоциации факторов транскрипции с РНК-полимеразой либо напрямую, либо через белки-посредники. Полимераза ассоциирует сначала с белками, а потом начинает транскрипцию[78]. В других случаях факторы транскрипции могут присоединяться к ферментам, которые модифицируют находящиеся на промоторах гистоны, что изменяет доступность ДНК для полимераз[79].
Так как специфические последовательности встречаются во многих местах генома, изменения в активности одного типа фактора транскрипции могут изменить активность тысяч генов[80]. Соответственно, эти белки часто регулируются в процессах ответа на изменения в окружающей среде, развития организма и дифференцировки клеток. Специфичность взаимодействия факторов транскрипции с ДНК обеспечивается многочисленными контактами между аминокислотами и основаниями ДНК, что позволяет им «читать» последовательность ДНК. Большинство контактов с основаниями происходит в главной бороздке, где основания более доступны[25].
Ферменты, модифицирующие ДНК[править |
Рибонуклеиновые кислоты — это… Что такое Рибонуклеиновые кислоты?
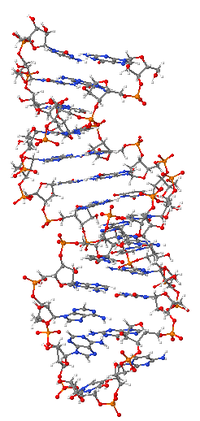
Рибонуклеи́новые кисло́ты (РНК) — нуклеиновые кислоты, полимеры нуклеотидов, в состав которых входят остаток ортофосфорной кислоты, рибоза (в отличие от ДНК, содержащей дезоксирибозу) и азотистые основания — аденин, цитозин, гуанин и урацил (в отличие от ДНК, содержащей вместо урацила тимин). Эти молекулы содержатся в клетках всех живых организмов, а также в некоторых вирусах.
Клеточные РНК образуются в ходе процесса, называемого транскрипцией, то есть синтеза РНК на матрице ДНК, осуществляемого специальными ферментами — РНК-полимеразами. Затем матричные РНК (мРНК) подвергаются сплайсингу и принимают участие в процессе, называемом трансляцией. Трансляция — это синтез белка на матрице мРНК при участии рибосом. Другие РНК после транскрипции подвергаются химическим модификациям, и после образования вторичной и третичной структур выполняют функции, зависящие от типа РНК.
Для одноцепочечных РНК характерны разнообразные пространственные структуры, в которых часть нуклеотидов одной и той же цепи спарены между собой. Некоторые высокоструктурированные РНК принимают участие в синтезе белка клетки, например, транспортные РНК служат для узнавания кодонов и доставки соответствующих аминокислот к месту синтеза белка, а рибосомные РНК служат структурной и каталитической основой рибосом.
Однако функции РНК в современных клетках не ограничиваются их ролью в трансляции. Так малые ядерные РНК принимают участие в сплайсинге эукариотических матричных РНК и других процессах.
Помимо того, что молекулы РНК входят в состав некоторых ферментов (например, теломеразы) у отдельных РНК обнаружена собственная энзиматическая активность, способность вносить разрывы в другие молекулы РНК или, наоборот, «склеивать» два РНК-фрагмента. Такие РНК называются рибозимами.
Геномы ряда вирусов состоят из РНК, то есть у них она играет роль, которую у высших организмов выполняет ДНК. На основании разнообразия функций РНК в клетке была выдвинута гипотеза, согласно которой РНК — первая молекула, которая была способна к самовоспроизведению в добиологических системах.
История изучения
Нуклеиновые кислоты были открыты в 1868 году швейцарским учёным Иоганном Фридрихом Мишером, который назвал эти вещества «нуклеин», поскольку они были обнаружены в ядре (лат. nucleus)[1]. Позже было обнаружено, что бактериальные клетки, в которых нет ядра, тоже содержат нуклеиновые кислоты. Значение РНК в синтезе белков было предположено в 1939 году в работе Торбьёрна Оскара Касперссона, Жана Брачета и Джека Шульца [2]. Джерард Маирбакс выделил первую матричную РНК, кодирующую гемоглобин кролика и показал, что при её введении в ооциты образуется тот же самый белок[3]. В Советском Союзе в 1956-57 годах проводились работы (А. Белозёрский, А. Спирин, Э. Волкин, Ф. Астрахан) по определению состава РНК клеток, которые привели к выводу, что основную массу РНК в клетке составляет рибосомальная РНК.[4] Северо Очоа получил Нобелевскую премию по медицине в 1959 году за открытие механизма синтеза РНК[5]. Последовательность 77 нуклеотидов одной из тРНК дрожжей S. cerevisiae была определена в 1965 году в лаборатории Роберта Холея, за что в 1968 году он получил Нобелевскую премию по медицине [6]. В 1967 Карл Вёзе предположил, что РНК обладают каталитическими свойствами. Он выдвинул так называемую Гипотезу РНК-мира, в котором РНК прото-организмов служила и в качестве молекулы хранения информации (сейчас эта роль выполняется в основном ДНК) и молекулы, которая катализировала метаболические реакции (сейчас это делают в основном ферменты)[7]. В 1976 Уолтер Фаэрс и его группа в Гентском Университете (Голландия) определили первую последовательность генома РНК-содержащего вируса, бактериофага MS2[8]. В начале 1990-х было обнаружено, что введение чужеродных генов в геном растений приводит к подавлению выражения аналогичных генов растения[9]. Приблизительно в это же время было показано, что РНК длиной около 22 оснований, которые сейчас называются микроРНК, играют регуляторную роль в онтогенезе нематод C.elegans [10].
Химический состав и модификации мономеров
Химическое строение полинуклеотида РНКНуклеотиды РНК состоят из сахара — рибозы, к которой в положении 1′ присоединено одно из оснований: аденин, гуанин, цитозин или урацил. Фосфатная группа соединяет рибозы в цепочку, образуя связи с 3′ атомом углерода одной рибозы и в 5′ положении другой. Фосфатные группы при физиологическом рН отрицательно заряжены, поэтому РНК — полианион. РНК транскрибируется как полимер четырёх оснований (аденина (A), гуанина (G), урацила (U) и цитозина (C), но в «зрелой» РНК есть много модифицированных оснований и сахаров[11]. Всего в РНК насчитывается около 100 разных видов модифицированных нуклеозидов, из которых 2′-О-метилрибоза наиболее частая модификация сахара, а псевдоуридин — наиболее часто встречающееся модифицированное основание [12]. У псевдоуридина (Ψ) связь между урацилом и рибозой не C — N, а C — C, этот нуклеотид встречается в разных положениях в молекулах РНК. В частности, псевдоуридин важен для функционирования тРНК[13]. Другое заслуживающее внимания модифицированное основание — гипоксантин, деаминированный гуанин, нуклеозид которого носит название инозина. Инозин играет важную роль в обеспечении вырожденности генетического кода. Роль многих других модификаций не до конца изучена, но в рибосомальной РНК многие пост-транскрипционные модификации находятся в важных для функционирования рибосомы участках. Например, на одном из рибонуклеотидов, участвующим в образовании пептидной связи[14].
Структура
Азотистые основания в составе РНК могут образовывать водородные связи между цитозином и гуанином, аденином и урацилом, а также между гуанином и урацилом [15]. Однако возможны и другие взаимодействия, например, несколько аденинов могут образовывать петлю, или петля, состоящая из четырёх нуклеотидов, в которой есть пара оснований аденин — гуанин[16].
Разные формы нуклеиновых кислот. На рисунке (слева направо) представлены A (типична для РНК), B (ДНК) и Z (редкая форма ДНК)Важная структурная особенность РНК, отличающая её от ДНК — наличие гидроксильной группы в 2′ положении рибозы, которая позволяет молекуле РНК существовать в А, а не В-конформации, наиболее часто наблюдаемой у ДНК[17]. У А-формы глубокая и узкая большая бороздка и неглубокая и широкая малая бороздка[18]. Второе последствие наличия 2′ гидроксильной группы состоит в том, что конформационно пластичные, то есть не принимающие участие в образовании двойной спирали, участки молекулы РНК могут химически атаковать другие фосфатные связи и их расщеплять [19].
«Рабочая» форма одноцепочечной молекулы РНК, как и у белков, часто обладает третичной структурой. Третичная структура образуется на основе элементов вторичной структуры, образуемой с помощью водородных связей внутри одной молекулы. Различают несколько типов элементов вторичной структуры — стебель-петли, петли и псевдоузлы[20]. В силу большого числа возможных вариантов спаривания оснований предсказание вторичной структуры РНК — гораздо более сложная задача, чем предсказание вторичной структуры белков, но в настоящее время есть эффективные программы, например, mfold [21].
Примером зависимости функции молекул РНК от их вторичной структуры являются участки внутренней посадки рибосомы (IRES). IRES — структура на 5′ конце информационной РНК, которая обеспечивает присоединение рибосомы в обход обычного механизма инициации синтеза белка, требующего наличия особого модифицированного основания (кэпа) на 5′ конце и белковых факторов инициации. Первоначально IRES были обнаружены в вирусных РНК, но сейчас накапливается всё больше данных о том, что клеточные мРНК также используют IRES-зависимый механизм инициации в условиях стресса[22].
Многие типы РНК, например, рРНК и мяРНК в клетке функционируют в виде комплексов с белками, которые ассоциируют с молекулами РНК после их синтеза или (у эукариот) экспорта из ядра в цитоплазму. Такие РНК-белковые комплексы называются рибонуклеопротеиновыми комплексами или рибонуклеопротеидами.
Сравнение с ДНК
Между ДНК и РНК есть три основных отличия:
- ДНК содержит сахар дезоксирибозу, РНК — рибозу, у которой есть дополнительная, по сравнению с дезоксирибозой, гидроксильная группа. Эта группа увеличивает вероятность гидролиза молекулы, то есть уменьшает стабильность молекулы РНК.
- Нуклеотид, комплементарный аденину, в РНК не тимин, как в ДНК, а урацил — неметилированная форма тимина.
- ДНК существует в форме двойной спирали, состоящей из двух отдельных молекул. Молекулы РНК, в среднем, гораздо короче и преимущественно одноцепочечные.
Структурный анализ биологически активных молекул РНК, включая тРНК, рРНК, мяРНК и другие молекулы, которые не кодируют белков, показал, что они состоят не из одной длинной спирали, а из многочисленных коротких спиралей, расположенных близко друг к другу и образующих нечто, похожее на третичную структуру белка. В результате этого РНК может катализировать химические реакции, например, пептидил-трансферазный центр рибосомы, участвующий в образовании пептидной связи белков, полностью состоит из РНК [23][24].
Синтез
Синтез РНК в живой клетке проводится ферментом — РНК-полимеразой. У эукариот разные типы РНК синтезируются разными, специализированными РНК-полимеразами. В целом матрицей синтеза РНК может выступать как ДНК, так и другая молекула РНК. Например, полиовирусы используют РНК-зависимую РНК-полимеразу для репликации своего генетического материала, состоящего из РНК[25]. Но РНК-зависимый синтез РНК, который раньше считался характерным только для вирусов, происходит и в клеточных организмах, в процессе так называемой РНК-интерференции[26].
Как в случае ДНК-зависимой РНК-полимеразы, так и в случае РНК-зависимой РНК-полимеразы фермент присоединяется к промоторной последовательности. Вторичная структура молекулы матрицы расплетается с помощью хеликазной активности полимеразы, которая при движении субстрата в направлении от 5′ к 3′ концу молекулы синтезирует РНК в 3′ —> 5′ направлении. Терминатор транскрипции в исходной молекуле определяет окончание синтеза. Многие молекулы РНК синтезируются в качестве молекул-предшественников, которые подвергаются «редактированию» — удалению ненужных частей с помощью РНК-белковых комплексов[27].
Например, у кишечной палочки гены рРНК расположены в составе одного оперона (в rrnB порядок расположения такой: 16S — tRNAGlu2 — 23S —5S) считываются в виде одной длинной молекулы, которая затем подвергается расщеплению в нескольких участках с образованием сначала пре-рРНК, а затем зрелых молекул рРНК[28]. Процесс изменения нуклеотидной последовательности РНК после синтеза носит название процессинга или редактирования РНК.
После завершения транскрипции РНК часто подвергается модификациям (см. выше), которые зависят от функции, выполняемой данной молекулой. У эукариот процесс «созревания» РНК, то есть её подготовки к синтезу белка, часто включает сплайсинг: удаление некодирующих белок последовательностей (интронов) с помощью рибонуклеопротеида сплайсосомы. Затем к 5′ концу молекулы пре-мРНК эукариот добавляется особый модифицированный нуклеотид (кэп), а к 3′ концу несколько аденинов, так назваемый «полиА-хвост»[27].
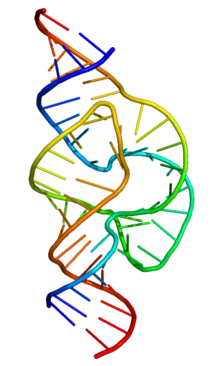 Структура молоточкового (hammerhead) рибозима, который расщепляет РНК
Структура молоточкового (hammerhead) рибозима, который расщепляет РНК.
Типы РНК
Матричная (информационная) РНК — РНК, которая служит посредником при передаче информации, закодированной в ДНК к рибосомам, молекулярным машинам, синтезирующим белки живого организма. Кодирующая последовательность мРНК определяет последовательность аминокислот полипептидной цепи белка [29]. Однако подавляющее большинство РНК не кодируют белок. Эти некодирующие РНК могут транскрибироваться с отдельных генов (например, рибосомальные РНК) или быть производными интронов [30]. Классические, хорошо изученные типы некодирующих РНК — это транспортные РНК (тРНК) и рРНК, которые участвуют в процессе трансляции [31]. Существуют также классы РНК, ответственные за регуляцию генов, процессинг мРНК и другие роли. Кроме того, есть и молекулы некодирующих РНК, способные катализировать химические реакции, такие, как разрезание и лигирование молекул РНК[32]. По аналогии с белками, способными катализировать химические реакции — энзимами (ферментами), каталитические молекулы РНК называются рибозимами.
Участвующие в трансляции
Информация о последовательности аминокислот белка содержится в мРНК. Три последовательных нуклеотида (кодон) соответствуют одной аминокислоте. В эукариотических клетках транскирибированный предшественник мРНК или пре-мРНК процессируется с образованием зрелой мРНК. Процессинг включает удаление некодирующих белок последовательностей (интронов). После этого мРНК экспортируется из ядра в цитоплазму, где к ней присоединяются рибосомы, транслирующие мРНК с помощью соединённых с аминокислотами тРНК.
В безъядерных клетках (бактерии и археи) рибосомы могут присоединяться к мРНК сразу после транскрипции участка РНК. И у эукариот, и у прокариот цикл жизни мРНК завершается её контролируемым разрушением ферментами рибонуклеазами [29].
Транспортные (тРНК) — малые, состоящие из приблизительно 80 нуклеотидов, молекулы с консервативной третичной структурой. Они переносят специфические аминокислоты в место синтеза пептидной связи в рибосоме. Каждая тРНК содержит участок для присоединения аминокислоты и антикодон для узнавания и присоединения к кодонам мРНК. Антикодон образует водородные связи с кодоном, что помещает тРНК в положение, способствующее образованию пептидной связи между последней аминокислотой образованного пептида и аминокислотой, присоединённой к тРНК [30].
Рибосомальные РНК (рРНК) — каталитическая составляющая рибосом. Эукариотические рибосомы содержат четыре типа молекул рРНК: 18S, 5.8S, 28S и 5S. Три из четырёх типов рРНК синтезируются в ядрышке. В цитоплазме рибосомальные РНК соединяются с рибосомальными белками и формируют нуклеопротеин, называемый рибосомой[29]. Рибосома присоединяется к мРНК и синтезирует белок. рРНК составляет до 80 % РНК, обнаруживаемой в цитоплазме эукариотической клетки [33].
Необычный тип РНК, который действует в качестве тРНК и мРНК (тмРНК) обнаружен во многих бактериях и пластидах. При остановке рибосомы на дефектных мРНК без стоп-кодонов тмРНК присоединяет небольшой пептид, направляющий белок на деградацию [34].
Участвующие в регуляции генов
В живых клетках обнаружено несколько типов РНК, которые могут уменьшать степень выражения гена при комплементарности мРНК или самому гену. Микро-РНК (21-22 нуклеотида в длину) найдены у эукариот и оказывают воздействие через механизм РНК-интерференции. При этом комплекс микро-РНК и ферментов может приводить к метилированию нуклеотидов в ДНК промотора гена, что служит сигналом для уменьшения активности гена. При использовании другого типа регуляции мРНК, комплементарная микро-РНК, деградируется.[35]. Однако есть и миРНК, которые увеличивают, а не уменьшают экспрессию генов.[36]. Малые интерферирующие РНК (миРНК, 20-25 нуклеотидов) часто образуются в результате расщепления вирусных РНК, но существуют и эндогенные клеточные миРНК[37]. Малые интерферирующие РНК также действуют через РНК-интерференцию по сходным с микро-РНК механизмам[38]. У животных найдены так называемыме РНК, взаимодействующие с Piwi (piRNA, 29-30 нуклеотидов), действующие в половых клетках против транспозиции и играющие роль в образовании гамет[39][40]. Кроме того, piRNA могут эпигенетически наследоваться по материнской линии, передавая потомству своё свойство ингибировать экспрессию транспозонов[41].
Антисмысловые РНК широко распространены у бактерий, многие из них подавляют выражение генов, но некоторые активируют экспрессию[42]. Действуют антисмысловые РНК, присоединяясь к мРНК, что приводит к образованию двуцепочечных молекул РНК, которые деградируются ферментами.[43]. У эукариот обнаружены высокомолекулярные, мРНК-подобные молекулы РНК. Эти молекулы также регулируют выражение генов,[44]. В качестве примера можно привести Xist, присоединяющуюся и инактивирующую одну из двух Х-хромосом у самок млекопитающих.[45].
Кроме роли отдельных молекул в регуляции генов, регуляторные элементы могут формироваться в 5′ и 3′ нетранслируемых участках мРНК. Эти элементы могут действовать самостоятельно, предотвращая инициацию трансляции, либо присоединять белки, например, ферритин или малые молекулы, например, биотин.[46].
В процессинге РНК
Многие РНК принимают участие в модификации других РНК. Интроны вырезаются из пре-мРНК сплайсосомами, которые, кроме белков, содержат несколько малых ядерных РНК (мяРНК)[31]. Кроме того, интроны могут катализировать собственное вырезание[47]. Синтезированая в результате транскрипции РНК также может быть химически модифицирована. У эукариот химические модификации нуклеотидов РНК, например, их метилирование, выполняется малыми ядерными РНК (мяРНК, 60-300 нуклеотидов). Этот тип РНК локализуется в ядрышко и тельцах Кахаля [30]. После ассоциации мяРНК с ферментами, мяРНК связываются с РНК-мишенью путём образования пар между основаниями двух молекул, а ферменты модифицируют нуклеотиды РНК-мишени. Рибосомальные и транспортные РНК содержат много подобных модификаций, конкретное положение которых часто сохраняется в процессе эволюции. Также могут быть модифицированы мяРНК и сами мяРНК[48][49]Гидовые РНК осуществляют процесс редактирования РНК в кинетопласте — особом участке митохондрии протистов-кинетопластид (например, трипаносом).
Геномы, состоящие из РНК
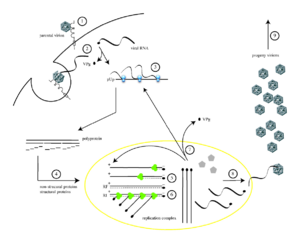 Жизненный цикл вируса с РНК геномом на примере полиовируса: 1 — присоединение исходного вириона к рецептору; 2 — вирион попадает в клетку; 3 — трансляция белков вируса с его РНК с образованием полипетида; 4 — полимеразы вируса размножают его РНК
Жизненный цикл вируса с РНК геномом на примере полиовируса: 1 — присоединение исходного вириона к рецептору; 2 — вирион попадает в клетку; 3 — трансляция белков вируса с его РНК с образованием полипетида; 4 — полимеразы вируса размножают его РНККак и ДНК, РНК может хранить информацию о биологических процессах. РНК может использоваться в качестве генома вирусов и вирусоподобных частиц. РНК-геномы можно разделить на те, которые не имеют промежуточной стадии ДНК и те, которые для размножения копируются в ДНК-копию и обратно в РНК (ретровирусы).
РНК-содержащие вирусы
Многие вирусы, например, вирус гриппа, на всех стадиях содержат геном, состоящий исключительно из РНК. РНК содержится внутри обычно белковой оболочки и реплицируется с помощью закодированных в ней РНК-зависимых РНК-полимераз. Вирусные геномы, состоящие из РНК разделяются на
- содержащие «плюс-цепь РНК», которая используется в качестве и мРНК, и генома;
- «минус-цепь РНК», которая служит только геномом, а в качестве мРНК используется комплементарная ей молекула;
- двухцепоченые вирусы.
Вироиды — другая группа патогенов, содержащих РНК-геном и не содержащих белок. Они реплицируются РНК-полимеразами организма хозяина[50].
Ретровирусы и ретротранспозоны
У других вирусов РНК-геном есть в течение только одной из фаз жизненного цикла. Вирионы так называемых ретровирусов содержат молекулы РНК, которые при попадании в клетки хозяина служат матрицей для синтеза ДНК-копии. В свою очередь, с матрицы ДНК считывается РНК-геном. Кроме вирусов обратную транскрипции применяют и класс мобильных элементов генома — ретротранспозоны [51]
Гипотеза РНК-мира
Способность молекул РНК одновременно служить как в качестве носителя информации, так и в качестве катализатора химических реакций, позволила выдвинуть гипотезу о том, что РНК была первым сложным полимером, появившимся в процессе добиологической эволюции. Эта гипотеза названа «гипотеза РНК-мира» [52][53]. Согласно ей, РНК на первых этапах эволюции автокатализировала синтез других молекул РНК, а затем и ДНК. На втором этапе эволюции синтезированные молекулы ДНК, как более стабильные, стали хранилищем генетической информации. Синтез белка на матрице РНК с помощью пра-рибосом, полностью состоящих из РНК, расширил свойства добиологических систем, постепенно белок заменил РНК в структурных аспектах. Из этой гипотезы делается вывод, что многие РНК, принимающие участие в синтезе белка в современных клетках, в особенности рРНК и тРНК — это реликты РНК-мира.
См. также
Примечания
Рекомендуемая литература
Ссылки
| Типы нуклеиновых кислот | |
|---|---|
| Азотистые основания | Пурины (Аденин, Гуанин) | Пиримидины (Урацил, Тимин, Цитозин) |
| Нуклеозиды | Аденозин | Гуанозин | Уридин | Тимидин | Цитидин |
| Нуклеотиды | монофосфаты (АМФ, ГМФ, UMP, ЦМФ) | дифосфаты (АДФ, ГДФ, УДФ, ЦДФ) | трифосфаты (АТФ, ГТФ, УТФ, ЦТФ) | циклические (цАМФ, цГМФ, cADPR) |
| Рибонуклеиновые кислоты | РНК | мРНК | тРНК | рРНК | антисмысловые РНК | gRNA | микроРНК | некодирующие РНК | piwi-interacting RNA | shRNA | малые интерферирующие РНК | малые ядерные РНК | малые ядрышковые РНК | тмРНК |
| Дезоксирибонуклеиновые кислоты | ДНК | кДНК | Геном | msDNA | Митохондриальная ДНК |
| Аналоги нуклеиновых кислот | en:glycerol nucleic acid | en:locked nucleic acid | ПНК | ТНК | Морфолино |
| Типы векторов | en:phagemid | Плазмиды | Фаг лямбда | en:cosmid | en:P1 phage | en:fosmid | en:Bacterial artificial chromosome | en:Yeast artificial chromosome | en:Human artificial chromosome |
Что значит рнк лабильна
РНК (рибонуклеиновая кислота), так же как и ДНК, относится к нуклеиновым кислотам. Молекулы-полимеры РНК намного меньше, чем у ДНК. Однако в зависимости от типа РНК количество входящих в них нуклеотидов-мономеров различается.
В состав нуклеотида РНК в качестве сахара входит рибоза, в качестве азотистого основания — аденит, гуанин, урацил, цитозин. Урацил по строению и химическим свойствам близок к тимину, который обычен для ДНК. В зрелых молекулах РНК многие азотистые основания модифицированы, поэтому в реальности разновидностей азотистых оснований в составе РНК намного больше.
Рибоза в отличие от дезоксирибозы имеет дополнительную -ОН-группу (гидроксильную). Это обстоятельство позволяет РНК легче вступать в химические реакции.
Главной функцией РНК в клетках живых организмов можно назвать реализацию генетической информации. Именно благодаря разным типам рибонуклеиновой кислоты генетический код считывается (транскрибируется) с ДНК, после чего на его основе синтезируются полипептиды (происходит трансляция). Итак, если ДНК в основном отвечает за хранение и передачу из поколения в поколение генетической информации (основной процесс – репликация), то РНК реализует эту информацию (процессы транскрипции и трансляции). При этом транскрипция происходит на ДНК, так что этот процесс относится к обоим типам нуклеиновых кислот и тогда с этой точки зрения можно сказать, что и ДНК отвечает за реализацию генетической информации.
При более подробном рассмотрении функции РНК намного разнообразнее. Ряд молекул РНК выполняют структурную, каталитическую и другие функции.
Существует так называемая гипотеза РНК-мира, согласно которой вначале в живой природе в качестве носителя генетической информации выступали только молекулы РНК, при этом другие молекулы РНК катализировали различные реакции. Данная гипотеза подтверждена рядом опытов, показывающих возможную эволюцию РНК. На это указывает и то, что ряд вирусов в качестве нуклеиновой кислоты, хранящей генетическую информацию, имеют молекулу РНК.
Согласно гипотезе РНК-мира ДНК появилась позже в процессе естественного отбора как более устойчивая молекула, что важно для хранения генетической информации.
Выделяют три основных типа РНК (кроме них есть и другие): матричная (она же информационная), рибосомальная и транспортная. Обозначаются они соответственно иРНК (или мРНК), рРНК, тРНК.
Информационная РНК (иРНК)
Почти все РНК синтезируются на ДНК в процессе транскрипции. Однако часто транскрипция упоминается как синтез именно информационной РНК (иРНК). Связано это с тем, что последовательность нуклеотидов иРНК в последствии определит последовательность аминокислот синтезируемого в процессе трансляции белка.
Перед транскрипцией нити ДНК расплетаются, и на одной из них с помощью комплекса белков-ферментов синтезируется РНК по принципу комплементарности, так же как это происходит при репликации ДНК. Только напротив аденина ДНК к молекуле РНК присоединяется нуклеотид, содержащий урацил, а не тимин.
На самом деле на ДНК синтезируется не готовая информационная РНК, а ее предшественник — пре-иРНК. Предшественник содержит участки последовательности нуклеотидов, которые не кодируют белок и которые после синтеза пре-иРНК вырезаются при участии малых ядерных и ядрышковых РНК («дополнительные» типы РНК). Эти удаляющиеся участки называются интронами. Остающиеся части иРНК называются экзонами. После удаления интронов экзоны сшиваются между собой. Процесс удаления интронов и сшивания экзонов называется сплайсингом. Усложняющей жизнь особенностью является то, что можно вырезать интроны по-разному, в результате получатся разные готовые иРНК, которые будут служить матрицами для разных белков. Таким образом, вроде бы один ген ДНК может играть роль нескольких генов.
Следует отметить, что у прокариотических организмов сплайсинга не происходит. Обычно их иРНК сразу после синтеза на ДНК готова к трансляции. Бывает, что пока конец молекулы иРНК еще транскрибируется, на ее начале уже сидят рибосомы, синтезирующие белок.
После того как пре-иРНК созревает в информационную РНК и оказывается вне ядра, она становится матрицей для синтеза полипептида. При этом на нее «насаживаются» рибосомы (не сразу, какая-то оказывается первой, другая — второй и т. д.). Каждая синтезирует свою копию белка, т. е. на одной молекуле РНК могут синтезироваться сразу несколько одинаковых белковых молекул (понятно, что каждая будет находиться на своей стадии синтеза).
Рибосома, передвигаясь от начала иРНК к ее концу, считывает по три нуклеотида (хотя вмещает шесть, т. е. два кодона) и присоединяет соответствующую транспортную РНК (имеющую соответствующий кодону антикодон), к которой присоединена соответствующая аминокислота. После этого с помощью активного центра рибосомы ранее синтезированная часть полипептида, соединенная с предшествующей тРНК, как-бы «пересаживается» (образуется пептидная связь) на аминокислоту, прикрепленную к только что пришедшей тРНК. Таким образом, молекула белка постепенно увеличивается.
Когда молекула информационной РНК становится не нужна, клетка ее разрушает.
Транспортная РНК (тРНК)
Транспортная РНК — это достаточно маленькая (по меркам полимеров) молекула (количество нуклеотидов бывает разным, в среднем около 80-ти), во вторичной структуре имеет форму клеверного листа, в третичной сворачивается в нечто подобное букве Г.
Функция тРНК — присоединение к себе соответствующей своему антикодону аминокислоты. В дальнейшем соединение с рибосомой, находящейся на соответствующем антикодону кодоне иРНК, и «передача» этой аминокислоты. Обобщая, можно сказать, что транспортная РНК переносит (на то она и транспортная) аминокислоты к месту синтеза белка.
Живая природа на Земле использует всего около 20-ти аминокислот для синтеза различных белковых молекул (на самом деле аминокислот куда больше). Но поскольку, согласно генетическому коду, кодонов больше 60-ти, то каждой аминокислоте может соответствовать несколько кодонов (на самом деле какой-то больше, какой-то меньше). Таким образом, разновидностей тРНК больше 20, при этом разные транспортные РНК переносят одинаковые аминокислоты. (Но и тут не так все просто.)
Рибосомная РНК (рРНК)
Рибосомную РНК часто также называют рибосомальной РНК. Это одно и то же.
Рибосомная РНК составляет около 80% всей РНК клетки, так как входит в состав рибосом, коих в клетке бывает достаточно много.
В рибосомах рРНК образует комплексы с белками, выполняет структурную и каталитическую функции.
В состав рибосомы входят несколько разных молекул рРНК, отличающиеся между собой как по длине цепи, вторичной и третичной структуре, выполняемым функциям. Однако их суммарная функция — это реализация процесса трансляции. При этом молекулы рРНК считывают информацию с иРНК и катализируют образование пептидной связи между аминокислотами.
| Показатели. | ДНК | РНК | АТФ |
| Нахождение в клетке | Ядро, митохондрии, пластиды. | Ядро, рибосомы, митохондрии, хлоропласты. | Цитоплазма, ядро, митохондрии. хлоропласты. |
| Нахождение в ядре. | Хроматин, хромосомы. | Ядрышко. | Кариоплазма. |
| Строение. | Две длинные полинуклеотидные цепочки, спирально закрученные антипараллельно относительно друг друга. | Одна короткая полинуклеотидная цепочка. | Мононуклеотид. |
| Мономеры. | Дезоксирибонуклеотиды. | Рибонуклеотиды. | Нет |
| Состав нуклеотида. | 1) азотистое основание — А, Г, Ц, Т, 2)углевод — дезоксирибоза 3)остаток фосфорной кислоты | 1)азотистое основание — А, Г, Ц, У, 2)углевод — рибоза 3) остаток фосфорной кислоты | 1)азотистое основание — А, 2)углевод 1 рибоза 3)три остатка фосфорной кислоты |
| Типы нуклеотидов. | Адениловый (А) Гуаниловый (Г) Цитидиловый (Ц) Тимидиловый (Т) | Адениловый (А) Гуаниловый (Г) Цитидиловый (Ц) Урациловый (У) | Адениловый (А) |
| Свойства. | 1) Способна к редуплекации или репликации (удвоению) по принципу комплеметарности (взаимодополняемость или соответствие) т.е. образование водородных святей между А-Т, Г-Ц, 2) Стабильна (не меняет место нахождения). | 1)Неспособна к редуплекации, кроме РНК вирусов, 2) Лабильна (переходит из ядра в цитоплазму). | В результате гидролиза от АТФ по одному отщепляются остатки фосфорной кислоты и высвобождается энергия. АТФ-АДФ-АМФ |
| Функции. | 1) Хранит, передаёт и воспроизводит генетическую информацию 2) Регулирует жизнедеятельность клетки. | 1) Участвует в биосинтезе белка а) и-РНК и м-РНК переносят генетическую информацию от ДНК к месту синтеза белка, б) р-РНК образует рибосому, в) т-РНК находит и переносит аминокислоты к месту синтеза белка, 2) в-РНК хранит, передаёт и воспроизводит генетическую информацию вируса. | 1) Энергетическая. |
| Особенности. | 1) Ядерная ДНК длинная, связана с белками и образует линейную хромосому. 2) Митохондриальная короткая и кольцевая, связана с белками и образует кольцевую хромосому. 3) У прокариот ДНК замкнута в кольцо, не связана с белками и не образует хромосому. | 1) Двухцепочечные РНК встречаются у некоторых вирусов. 2) 5 видов РНК: и-РНК информационная. м-РНК матричная, р-РНК рибосомная, т-РНК транспортная, в-РНК вирусная | 1) Остатки фосфорной кислоты соединены между собой макроэргическими (высокоэнергетическими) связями. 2) Молекула АТФ неустойчива, существует менее 1 минуты, восстанавливается и расщепляется 2400 раз в сутки. |
Репликация ДНК, генетический код, реализация генетической информации.
3.1. Репликация ДНК. Поскольку ДНК является молекулой наследственности, то для реализации этого свойства она должна точно копировать саму себя и таким образом сохранять имеющуюся в исходной молекуле ДНК информацию в виде определённой последовательности нуклеотидов. Это обеспечивается за счёт особого процесса, который называется репликацией или редупликацией.
Репликация — это удвоение молекулы ДНК. В основе репликации лежат правила Эдвина Чаргаффа (А+Г=Т+Ц) т.е. сумма пуриновых оснований равна сумме пиримидиновых оснований. Строгое соответствие нуклеотидов друг другу в парных цепочках ДНК называется комплементарностью (взаимодополняемостью).
Этапы репликации:
| № | Этапы репликации. |
| Специальные ферменты раскручивают двойную спираль молекулы ДНК и разрывают водородные связи между цепочками. | |
| Фермент ДНК-полимераза движется вдоль одной цепочки ДНК от 3 атома углерода к 5 атому и по правилу комплементарности (А-Т, Г-Ц) присоединяет соответствующие нуклеотиды. Эта цепочка называется лидирующей, её удвоение идет непрерывно. | |
| Вторая цепочка отстающая расположена антипаралельно первой, а ДНК-полимераза 1 может двигаться только в одном направлении от З атома углерода к 5 атому, следовательно, она копируется отдельными фрагментами по мере раскручивания молекулы ДНК. Фрагменты сшиваются специальными ферментами — лигазами по принципу антипараллельности. | |
| После репликации каждая молекула ДНК содержит одну «материнскую» цепочку и вторую вновь синтезированную «дочернюю». Такой принцип синтеза называется полуконсервативным, т.е. одна цепочка в новой молекуле ДНК «старая», а вторая «новая». |
Генетический код.
Для молекулы наследственности, которой является ДНК, свойственно не только самоудвоение (репликация), но и кодирование информации с помощь определённой последовательности нуклеотидов. Известно, что ДНК состоит из четырёх видов нуклеотидов, то есть информация в ДНК записывается 4 буквами (А, Т, Г, Ц). Математические расчёты показывают, что
1. Если использовать 1 нуклеотид, то получим 4 разных сочетания, 4 2 =16), 16 3 =64), 64>20.
Таким образом, комбинации из 3 нуклеотидов будет достаточно, чтобы закодировать 20 аминокислот. Из 64 возможных триплетов 61 триплет кодирует 20 незаменимых аминокислот, обнаруженных в составе клеточных белков, а 3 триплета являются стоп- сигналами или терминаторами, которые прекращают считывание информации.
Сочетания из трёх нуклеотидов, кодирующие определённые аминокислоты, называются кодом ДНК, или генетическим кодом. В настоящее время генетический код полностью расшифрован, то есть известно, какие триплетные сочетания нуклеотидов кодируют 20 аминокислот. Пользуясь комбинацией, состоящей из трёх нуклеотидов, можно закодировать больше аминокислот, чем необходимо для кодирования 20 аминокислот. Оказалось, что каждая аминокислота может кодироваться несколькими триплетами, кроме метионина и триптофана. Аминокислоты входящие в состав природных белков могут относится к разным группам, заменимые кислоты (З), незаменимые (НЗ).
Генетический код — это система записи генетической информации в ДНК в виде определённой последовательности нуклеотидов (или способ записи последовательности аминокислот в белке с помощью нуклеотидов).
Генетический код обладает несколькими свойствами (7 свойств).
Рибонуклеиновая кислота представляет собой сополимер пуриновых и пиримидиновых рибонуклеотидов, соединенных друг с другом, как и в ДНК, -фосфодиэфирными мостиками (рис. 37.6). Хотя эти два вида нуклеиновых кислот имеют много общего, по ряду признаков они отличаются друг от друга.
1. У РНК углеводным остатком, к которому присоединены пуриновые или пиримидиновые основания и фосфатные группы, является рибоза, а не 2-дезоксирибоза (как у ДНК).
2. Пиримидиновые компоненты РНК отличаются от таковых у ДНК. В состав РНК, как и в состав ДНК, входят нуклеотиды аденина, гуанина и цитозина. В то же время РНК (за исключением некоторых специальных случаев, на которых мы остановимся ниже) не содержит тимина, его место в молекуле РНК занимает урацил.
3. РНК — одноцепочечная молекула (в отличие от ДНК, имеющей двухцепочечную структуру), однако при наличии в цепи РНК участков с комплементарной последовательностью (противоположной полярности) единичная цепь РНК способна сворачиваться с образованием так называемых «шпилек», структур, имеющих двухспиральные характеристики (рис. 37.7).
Рис. 37.6. Фрагмент молекулы рибонуклеиновой кислоты (РНК), в котором пуриновые и пиримидиновые основания— аденин (А), урацил (U), цитозин (С) и гуанин (-удерживаются фосфодиэфирным остовом, соединяющим рибозильные остатки, связанные N-гликозидной связью с соответствующими нуклеиновыми основаниями. Обратите внимание: цепь РНК обладает определенной направленностью, на которую указывают 5- и З-концевые фосфатные остатки.
4. Так как молекула РНК представляет собой одиночную цепь, комплементарную только одной из цепей ДНК, содержание в ней гуанина не обязательно равно содержанию цитозина, а содержание аденина не обязательно равно содержанию урацила.
5. РНК может быть гидролизована щелочью до 2, З-циклических диэфиров мононуклеотидов; в роли промежуточного продукта гидролиза выступает 2, У, 5-триэфир, который не образуется при щелочном гидролизе ДНК из-за отсутствия у последней 2-гидроксильных групп; щелочная лабильность РНК (сравнительно с ДНК) является полезным свойством как для диагностических, так и для аналитических целей.
Информация, содержащаяся в одноцепочечной РНК, реализуется в виде определенной последовательности пуриновых и пиримидиновых оснований (т. е. в первичной структуре) полимерной цепи. Эта последовательность комплементарна кодирующей цепи гена, с которой «считывается» РНК. Вследствие комплементарности молекула РНК способна специфически связываться (гибридизоваться) с кодирующей цепью, но не гибридизуется с некодирующей цепью ДНК. Последовательность РНК (за исключением замены Т на U) идентична последовательности некодирующей цепи гена (рис. 37.8).
Биологические функции РНК
Известно несколько видов РНК. Почти все они непосредственно вовлечены в процесс биосинтеза белка. Молекулы цитоплазматической РНК, выполняющие функции матриц белкового синтеза, называются матричными РНК (мРНК). Другой вид цитоплазматической РНК—рибосомная РНК (рРНК) — выполняет роль структурных компонентов рибосом (органелл, играющих важную роль в синтезе белка). Адапторные молекулы транспортных РНК (тРНК) участвуют в трансляции (переводе) информации мРНК в последовательность аминокислот в белках.
Значительная часть РНК—первичных транскриптов, образующихся в эукариотических клетках, включая и клетки млекопитающих, — подвергается деградации в ядре и не играет какой-либо структурной или информационной роли в цитоплазме. В культивируемых
Рис. 37.7. Вторичная структура молекулы РНК типа «петли со стеблем» («шпилька»), возникающая вследствие внутримолекулярного образования водородных связей между комплементарными парами нуклеиновых оснований.
клетках человека обнаружен класс малых ядерных РНК которые непосредственно не участвуют в синтезе белка, но могут оказывать влияние на процессинг РНК и общую «архитектуру» клетки. Размеры этих относительно небольших молекул варьируют, последние содержат от 90 до 300 нуклеотидов (табл. 37.3).
РНК является основным генетическим материалом у некоторых вирусов животных и растений. Некоторые РНК-содержащие вирусы никогда не проходят стадию обратной транскрипции РНК в ДНК. Однако для большинства известных вирусов животных, таких, как ретровирусы, характерна обратная транскрипция их РНК-генома, направляемая РНК-зависимой ДНК-полимеразой (обратной транскриптазой) с образованием двухспиральной ДНК-копии. Во многих случаях образующийся двухспиральный ДНК-транскрипт встраивается в геном и в дальнейшем обеспечивает экспрессию генов вируса, а также наработку новых копий вирусных РНК-геномов.
Структурная организация РНК
Во всех эукариотических и прокариотических организмах существуют три основных класса молекул РНК: информационная (матричная или мессенджер) РНК (мРНК), транспортная (тРНК) и рибосомная (рРНК). Представители этих классов отличаются друг от друга размерами, функциями и стабильностью.
Информационная (мРНК) — наиболее гетерогенный в отношении размеров и стабильности класс. Все представители этого класса служат переносчиками информации от гена к белок-синтезирующей системе клетки. Они выполняют роль матриц для синтезируемого полипептида, т. е. определяют аминокислотную последовательность белка (рис. 37.9).
Информационные РНК, особенно эукариотические, обладают некоторыми уникальными структурными особенностями. 5-Конец мРНК «кэпирован» 7-метилгуанозинтрифосфатом, присоединенным к 5-гидроксилу соседнего 2-0-метилрибонуклеозида через остаток трифосфата (рис. 37.10). Молекулы мРНК часто содержат внутренние остатки 6-метиладенина и 2-0-метилированные рибонуклеотиды. Хотя смысл «кэпирования» до конца еще не выяснен, можно предположить, что образующаяся структура 5-конца мРНК используется для специфического узнавания в системе трансляции. Синтез белка начинается на 5′-(кэпированном) конце мРНК. Другой конец большинства молекул мРНК (З-конец) содержит полиаденилатную цепочку из 20—250 нуклеотидов. Специфические функции этого окончательно не установлены. Можно предполагать, что данная структура отвечает за поддержание внутриклеточной стабильности мРНК. Некоторые мРНК, включая гистоновые не содержат poly (А). Наличие poly (А) в структуре мРНК используется для отделения от других видов РНК посредством фракционирования тотальной РНК на колонках с oligo (Т), иммобилизованным на твердом носителе типа целлюлозы. Связывание мРНК с колонкой происходит за счет комплементарных взаимодействий poly (А)-«хвоста» с иммобилизованным oligo (Т).
Рис. 37.8. Последовательность гена и его РНК-транскрипта. Показаны кодирующая и некодирующая цепи, и отмечена их полярность. РНК-транскрипт, имеющий полярность комплементарен кодирующей цепи (с полярностью 3 — 5) и идентичен по последовательности (за исключением замен Т на U) и полярности некодирующей цепи ДНК.
Рис. 37.9. Экспрессия генетической информации ДНК в форме мРНК-транскрипта и последующая трансляция при участии рибосом с образованием специфической молекулы белка.
Рис. 37.10. Структура «кэпа», находящегося на 5-конце большинства эукариотических матричных РНК 7-метилгуанозинтрифосфат присоединяется к 5-концу мРНК. на котором обычно находится 2-О-метилпуриновый нуклеотид.
В клетках млекопитающих, включая клетки человека, зрелые молекулы мРНК, находящиеся в цитоплазме, не являются полной копией транскрибируемого участка гена. Образующийся в результате транскрипции полирибонуклеотид представляет собой предшественник цитоплазматической мРНК, перед выходом из ядра он подвергается специфическому процессингу. Непроцессированные продукты транскрипции, обнаруживаемые в ядрах клеток млекопитающих, образуют четвертый класс молекул РНК. Такие ядерные РНК очень гетерогенны и достигают значительных размеров. Молекулы гетерогенных ядерных РНК могут иметь молекулярную массу более , в то время как молекулярная масса мРНК обычно не превышает 2106. подвергаются процессингу в ядре, и образующиеся зрелые мРНК поступают в цитоплазму, где служат матрицей для биосинтеза белка.
Молекулы транспортных РНК (тРНК) обычно содержат около 75 нуклеотидов. Молекулярная масса таких молекул составляет . тРНК также формируются в результате специфического процессинга соответствующих молекул-предшественников (см. гл. 39). Транспортные тРНК выполняют функцию посредников в ходе трансляции мРНК. В любой клетке присутствуют не менее 20 видов молекул тРНК. Каждый вид (иногда несколько видов) тРНК соответствует одной из 20 аминокислот, необходимых для синтеза белка. Хотя каждая специфическая тРНК отличается от других нуклеотидной последовательностью, все они имеют и общие черты. Благодаря нескольим внутрицепочечным комплементарным участкам, все тРНК обладают вторичной структурой, получившей название «клеверный лист» (рис. 37.11).
Молекулы всех видов тРНК имеют четыре основных плеча. Акцепторное плечо состоит из «стебля» спаренных нуклеотидов и заканчивается последовательностью ССА Именно через У-гидроксильную группу аденозильного остатка происходит связывание с карбоксильной группой аминокислоты. Остальные плечи тоже состоят из «стеблей», образованных комплементарными парами оснований, и петель из неспаренных оснований (рис. 37.7). Антикодоновое плечо узнает нуклеотидный триплет или кодон (см. гл. 40) в мРНК. D-плечо названо так из-за наличия в нем дигидроуридина, -плечо названо по последовательности Т-псевдоуридин-С. Дополнительное плечо представляет собой наиболее вариабельную структуру и служит основой классификации тРНК. тРНК класса 1 (75% от общего их числа) обладают дополнительным плечом длиной 3—5 пар оснований. Дополнительное плечо у тРНК-молекул класса 2 состоит из 13—21 пар оснований и часто включает неспаренную петлю.
Рис. 37.11. Структура молекулы аминоацил-тРНК, к 3-ССА-концу которой присоединена аминокислота . Указаны внутримолекулярные водородные связи и расположение антикодонового, ТТС- и дигидроурацилового плеч. (From J. D. Watson. Molecular biology of the Gene 3rd, ed.. Copyright 1976, 1970, 1965 by W. A. Benjamin, Inc., Menlo Park Calif.)
Вторичная структура, определяемая системой комплементарных взаимодействий нуклеотидных оснований соответствующих плеч, характерна для всех видов Акцепторное плечо содержит семь пар оснований, -плечо — пять пар оснований, плечо D — три (или четыре) пары оснований.
Молекулы тРНК весьма стабильны у прокариот и несколько менее стабильны у эукариот. Обратная ситуация характерна для мРНК, которая довольно нестабильна у прокариот, а у эукариотических организмов обладает значительной стабильностью.
Рибосомная РНК. Рибосома — это цитоплазматическая нуклеопротеиновая структура, предназначенная для синтеза белка по мРНК-матрице. Рибосома обеспечивает специфический контакт в результате которого и происходит трансляция нуклеотидной последовательности, считанной с определенного гена, в аминокислотную последовательность соответствующего белка.
В табл. 37.2 представлены компоненты рибосом млекопитающих, имеющих молекулярную массу 4,210 6 и скорость седиментации (единиц Сведберга). Рибосомы млекопитающих состоят из двух нуклеопротеиновых субъединиц — большой с
Таблица 37.2. Компоненты рибосом млекопитающих
молекулярной массой (60S), и малой, имеющей молекулярную массу (40S). 608-субъединица содержит 58-рибосомную РНК (рРНК), 5,8S-pPHK и 28S-pPHK, а также более 50 различных полипептидов. Малая, 408-субъединица включает единственную 18S-pPHK и около 30 полипептидных цепей. Все рибосомные РНК, за исключением 5S-PHK, имеют общего предшественника—45S-PHK, локализованную в ядрышке (см. гл. 40). У молекулы 5S-PHK предшественник собственный. В ядрышке происходит упаковка высокометилированных рибосомных РНК с рибосомными белками. В цитоплазме рибосомы достаточно устойчивы и способны осуществлять большое число циклов трансляции.
Небольшие стабильные РНК. В эукариотических клетках обнаружено большое число дискретных, высококонсервативных, небольших и стабильных молекул РНК. Большинство РНК этого типа обнаруживаются в составе рибонуклеопротеинов и локализованы в ядре, цитоплазме или одновременно в обоих компартментах. Размеры этих молекул варьируют от 90 до 300 нуклеотидов, содержание их — 100000—1000000 копий на клетку.
Малые ядерные нуклеопротеиновые частицы (часто называемые snurps — от англ. small nuclear ribonucleic particles), вероятно, играют существенную роль в регуляции экспрессии генов. Нуклеопротеиновые частицы типа U7, по-видимому, участвуют в формировании З-концов гистоновых мРНК. Частицы , вероятно, необходимы для полиаденилирования, a — для удаления интронов и процессинга мРНК (см. гл. 39). Табл. 37.3. суммирует некоторые характеристики небольших стабильных РНК.
Таблица 37.3. Некоторые виды небольших стабильных РНК, обнаруженные в клетках млекопитающих
ЛИТЕРАТУРА
Darnell J. et al. Molecular Cell Biology, Scientific American Books, 1986.
Hunt T. DNA Makes RNA Makes Protein, Elsevier, 1983. Lewin B. Genes, 2nd ed., Wiley, 1985.
Rich A. et al. The chemistry and biology of left-handed Z-DNA, Annu. Rev. Biochem., 1984, 53, 847.
Turner P. Controlling roles for snurps, Nature, 1985, 316, 105. Watson J. D. The Double Helix, Atheneum, 1968.
Watson J. D., Crick F.H.C. Molecular structure of nucleic acids. Nature, 1953, 171, 737.
Zieve G. W. Two groups of small stable RNAs, Cell, 1981, 25, 296.
Нуклеиновая кислота — Википедия
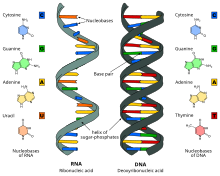 Нуклеиновые кислоты РНК (слева) и ДНК (справа).
Нуклеиновые кислоты РНК (слева) и ДНК (справа).Нуклеи́новая кислота (от лат. nucleus — ядро) — высокомолекулярное органическое соединение, биополимер (полинуклеотид), образованный остатками нуклеотидов. Нуклеиновые кислоты ДНК и РНК присутствуют в клетках всех живых организмов и выполняют важнейшие функции по хранению, передаче и реализации наследственной информации.
- В 1847 из экстракта мышц быка было выделено[1] вещество, которое получило название «инозиновая кислота». Это соединение стало первым изученным нуклеотидом. В течение последующих десятилетий были установлены детали его химического строения. В частности, было показано, что инозиновая кислота является рибозид-5′-фосфатом, и содержит N-гликозидную связь.
- В 1868 году швейцарским химиком Фридрихом Мишером при изучении некоторых биологических субстанций было открыто неизвестное ранее вещество. Вещество содержало фосфор и не разлагалось под действием протеолитических ферментов. Также оно обладало выраженными кислотными свойствами. Вещество было названо «нуклеином». Соединению была приписана брутто-формула C29H49N9O22P3.
- Уилсон обратил внимание на практическую идентичность химического состава «нуклеина» и открытого незадолго до этого «хроматина» — главного компонента хромосом[2]. Было выдвинуто предположение об особой роли «нуклеина» в передаче наследственной информации.
- В 1889 г Рихард Альтман ввел термин «нуклеиновая кислота», а также разработал удобный способ получения нуклеиновых кислот, не содержащих белковых примесей.
- Левин и Жакоб, изучая продукты щелочного гидролиза нуклеиновых кислот, выделили их основные составляющие — нуклеотиды и нуклеозиды, а также предложили структурные формулы, верно описывающие их химические свойства.
- В 1921 году Левин выдвинул гипотезу «тетрануклеотидной структуры ДНК»[3], оказавшуюся впоследствии ошибочной[4].
- В 1935 году Клейн и Танхаузер с помощью фермента фосфатазы провели мягкое фрагментирование ДНК, в результате чего были получены в кристаллическом состоянии четыре ДНК-образующих нуклеотида[5]. Это открыло новые возможности для установления структуры этих соединений.
- В 1940-е годы научная группа в Кембридже под руководством Александера Тодда проводит широкие синтетические исследования в области химии нуклеотидов и нуклеозидов. В результате их работы были установлены все детали химического строения и стереохимии нуклеотидов. За цикл работ в этой области Александер Тодд был награждён Нобелевской премией в области химии в 1957 году.
- В 1951 году Чаргаффом была установлена закономерность содержания в нуклеиновых кислотах нуклеотидов разных типов, получившая впоследствии название Правило Чаргаффа.
- В 1953 году Уотсоном и Криком установлена вторичная структура ДНК, двойная спираль[6].
 Гелеобразный осадок нуклеиновой кислоты
Гелеобразный осадок нуклеиновой кислотыОписаны многочисленные методики выделения нуклеиновых кислот из природных источников. Основными требованиями, предъявляемыми к методу выделения, являются эффективное отделение нуклеиновых кислот от белков, а также минимальная степень фрагментации полученных препаратов. Классический метод выделения ДНК был описан в 1952 году и используется в настоящее время без значительных изменений[7]. Клеточные стенки исследуемого биологического материала разрушаются одним из стандартных методов, а затем обрабатываются анионным детергентом. При этом белки выпадают в осадок, а нуклеиновые кислоты остаются в водном растворе. ДНК может быть осаждена в виде геля осторожным добавлением этанола к её солевому раствору. Концентрацию полученной нуклеиновой кислоты, а также наличие примесей (белки, фенол) обычно определяют спектрофотометрически по поглощению на А260 нм.
Нуклеиновые кислоты легко деградируют под действием особого класса ферментов — нуклеаз. В связи с этим при их выделении важно обработать лабораторное оборудование и материалы соответствующими ингибиторами. Так, например, при выделении РНК широко используется такой ингибитор рибонуклеаз как DEPC.
Нуклеиновые кислоты хорошо растворимы в воде, практически нерастворимы в органических растворителях. Очень чувствительны к действию температуры и критическим значениям уровня pH. Молекулы ДНК с высокой молекулярной массой, выделенные из природных источников, способны фрагментироваться под действием механических сил, например, при перемешивании раствора. Нуклеиновые кислоты фрагментируются ферментами — нуклеазами.
Фрагмент полимерной цепочки ДНКПолимерные формы нуклеиновых кислот называют полинуклеотидами.
Существуют 4 уровня структурной организации нуклеиновых кислот: первичная, вторичная, третичная и четвертичная. Первичная структура представляет собой цепочки из нуклеотидов, соединяющихся через остаток фосфорной кислоты (фосфодиэфирная связь). Вторичная структура — это две цепи нуклеиновых кислот соединённые водородными связями. Стоит отметить, что цепи соединяются по типу «голова-хвост» (3′ к 5′), по принципу комплементарности (азотистые основания находятся внутри этой структуры). Третичная структура, или же спираль, образуется за счет радикалов азотистых оснований (образуются водородные дополнительные связи, которые и сворачивают эту структуру, тем самым обуславливая её прочность). И наконец 4 структура — это комплексы гистонов и нитей хроматина.
Поскольку в нуклеотидах существует только два типа гетероциклических молекул, рибоза и дезоксирибоза, то и имеется лишь два вида нуклеиновых кислот — дезоксирибонуклеиновая (ДНК) и рибонуклеиновая (РНК).
Мономерные формы также встречаются в клетках и играют важную роль в процессах передачи сигналов или запасании энергии. Наиболее известный мономер РНК — АТФ, аденозинтрифосфорная кислота, важнейший аккумулятор энергии в клетке.
Ма́тричная рибонуклеи́новая кислота́ (мРНК, синоним — информацио́нная РНК, иРНК) — РНК, содержащая информацию о первичной структуре (аминокислотной последовательности) белков[8]. мРНК синтезируется на основе ДНК в ходе транскрипции, после чего, в свою очередь, используется в ходе трансляции как матрица для синтеза белков. Тем самым мРНК играет важную роль в «проявлении» (экспрессии) генов.
Рибосо́мные рибонуклеи́новые кисло́ты (рРНК) — несколько молекул РНК, составляющих основу рибосомы. Основной функцией рРНК является осуществление процесса трансляции — считывания информации с мРНК при помощи адапторных молекул тРНК и катализ образования пептидных связей между присоединёнными к тРНК аминокислотами.
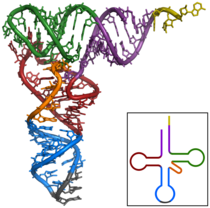 Структура транспортной РНК
Структура транспортной РНКТранспортная РНК, тРНК — рибонуклеиновая кислота, функцией которой является транспортировка аминокислот к месту синтеза белка. Имеет типичную длину от 73 до 93 нуклеотидов и размеры около 5 нм. тРНК также принимают непосредственное участие в наращивании полипептидной цепи, присоединяясь — будучи в комплексе с аминокислотой — к кодону мРНК и обеспечивая необходимую для образования новой пептидной связи конформацию комплекса.
Для каждой аминокислоты существует своя тРНК.
тРНК является одноцепочечной РНК, однако в функциональной форме имеет конформацию «клеверного листа». Аминокислота ковалентно присоединяется к 3′-концу молекулы с помощью специфичного для каждого типа тРНК фермента аминоацил-тРНК-синтетазы. На участке C находится антикодон, соответствующий аминокислоте.
Некодирующие РНК (non-coding RNA, ncRNA) — это молекулы РНК, которые не транслируются в белки. Ранее использовавшийся синоним, малые РНК (smRNA, small RNA), в настоящее время не используется, так как некоторые некодирующие РНК могут быть очень большими, например, Xist.
Последовательность ДНК, на которой транскрибируются некодирующие РНК, часто называют РНК-геном.
Некодирующие РНК включают в себя молекулы РНК, которые выполняют очень важные функции в клетке — транспортные РНК (тРНК), рибосомные РНК (рРНК), такие малые РНК, как малые ядрышковые РНК (snoRNA), микроРНК, siRNA, piRNA, а также длинные некодирующие РНК — Xist, Evf, Air, CTN, PINK, TUG1.
Последние транскриптомные технологии (секвенирование РНК) и методы ДНК-микрочипов предполагают наличие более 30000 длинных некодирующих РНК (англ. long ncRNA). Примерно такое же количество малых регуляторных РНК содержится в геноме мыши.
- ↑ J. Liebig. ??? (неопр.) // Annalen. — 1847. — Т. 62. — С. 257.
- ↑ Edmund B. Wilson. An Atlas of the Fertilization and Karyokinesis of the Ovum. — N. Y.: Macmillan, 1895. — P. 4.
- ↑ P. A. Levene. ??? (англ.) // J. Biol. Chem. : journal. — 1921. — Vol. 48. — P. 119.
- ↑ Во время выдвижения «тетрануклеотидной структуры» химики критически относились к самой возможности существования макромолекул, вследствие чего ДНК была приписана структура с низкой молекулярной массой
- ↑ W. Klein, S. J. Thannhauser. ??? (неопр.) // Z. physiol. Chem.. — 1935. — Т. 231. — С. 96.
- ↑ J. D. Watson, F. H. C. Crick. Molecular Structure of Nucleic Acids: A Structure for Deoxyribose Nucleic Acid (англ.) // Nature. — 1953. — Vol. 171. — P. 737—738. — doi:10.1038/171737a0.
- ↑ Ernest R. M. Kay, Norman S. Simmons, Alexander L. An Improved Preparation of Sodium Desoxyribonucleate (англ.) // J. Am. Chem. Soc. (англ.)русск. : journal. — 1952. — Vol. 74, no. 7. — P. 1724—1726. — doi:10.1021/ja01127a034.
- ↑ Bruce Alberts, Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, Peter Walter. Molecular Biology of the Cell. — 5. — Garland Science, 2008. — 1392 с. — ISBN 0815341059.
- Бартон Д., Оллис У. Д. Общая органическая химия. — М.: Химия, 1986. — Т. 10. — С. 32—215. — 704 с.
- Франк-Каменецкий М. Д. Самая главная молекула. — М.: Наука, 1983. — 160 с.
- Аппель Б., Бенеке И., Бенсон Я., под ред. С. Мюллер. Нуклеиновые кислоты от А до Я. — М.: Бином, 2012. — 352 с. — ISBN 978-5-9963-0376-2.
ИРНК — это… Что такое ИРНК?
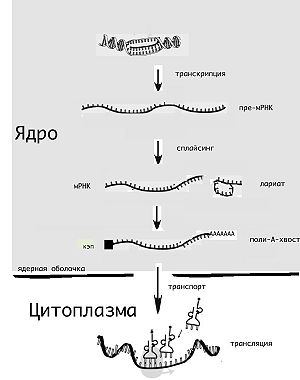
Основные этапы жизненного цикла мРНК эукариот
Ма́тричная рибонуклеи́новая кислота́ (мРНК, синоним — информацио́нная РНК, иРНК) — РНК, отвечающая за перенос информации о первичной структуре белков от ДНК к местам синтеза белков. мРНК синтезируется на основе ДНК в ходе транскрипции, после чего, в свою очередь, используется в ходе трансляции как матрица для синтеза белков. Тем самым мРНК играет важную роль в «проявлении» (экспрессии) генов.
Длина типичной зрелой мРНК составляет от нескольких сотен до нескольких тысяч нуклеотидов. Самые длинные мРНК отмечены у (+)оц РНК-содержащих вирусов, например пикорнавирусов, однако следует помнить, что у этих вирусов мРНК образует весь их геном.
ДНК нередко сравнивают с чертежами для изготовления белков. Развивая эту инженерно-производственную аналогию, можно сказать, что, если ДНК — это полный набор чертежей для изготовления белков, находящийся на хранении в сейфе директора завода, то мРНК — временная рабочая копия чертежа, выдаваемая в сборочный цех.
Гипотеза о значении РНК в синтезе белков была высказана Торбьёрном Касперссоном (Torbjörn Caspersson) на основе исследований 1937—1939 гг., в результате которых было показано, что клетки, активно синтезирующие белок, содержат большое количество РНК. Подтверждение гипотезы было получено Юбером Шантренне (Hubert Chantrenne).
«Жизненный цикл» мРНК
Жизненный цикл молекулы мРНК начинается её «считыванием» с матрицы ДНК (транскрипция) и завершается её деградацией до отдельных нуклеотидов. Молекула мРНК в течение своей жизни может быть также «отредактирована» перед синтезом белка (трансляцией). Эукариотические молекулы мРНК часто требуют сложной обработки и транспортировки из ядра — места синтеза мРНК, в цитоплазму, где происходит трансляция, в то время как прокариотические молекулы мРНК этого не требуют и синтез РНК у них сопряжён с синтезом белка.[1].
Транскрипция
Транскрипцией называют процесс копирования генетической информации с ДНК на РНК, в частности на мРНК. Транскрипция осуществляется ферментом РНК-полимеразой, строящей, согласно принципу комплементарности, копию участка ДНК на основании одной из цепей двойной спирали. Этот процесс как у эукариот, так и у прокариот организован одинаково. Основное различие между про- и эукариотами состоит в том, что у эукариот РНК-полимераза во время транскрипции ассоциируется с мРНК-обрабатывающими ферментами, поэтому у них обработка мРНК и транскрипция могут проходить одновременно. Короткоживущие необработанные или частично обработанные продукты транскрипции называются пред-мРНК; после полной обработки — зрелая мРНК.
Обработка эукариотической пред-мРНК
Основная статья: Посттранскрипционные изменения РНК
В то время как мРНК прокариот (бактерий и архей), за редкими исключениями, сразу готовы к трансляции и не требуют специальной обработки, эукариотические пре-мРНК требуют более интенсивной обработки. В процессе сплайсинга из пре-мРНК удаляются не кодирующие белок последовательности (интроны), на 5′ конец молекулы добавляется специальный модифицированный нуклеотид (кэп), на 3′ конец добавляются несколько аденинов, так называемый полиадениновый хвост. Кэп узнаётся факторами инициации, белками, отвечающими за присоединение к мРНК рибосомы, полиадениновый хвост связывается с со специальным белком. Обычно эти посттранскрипционные изменения мРНК эукариот обозначают термином «процессинг мРНК». Полиаденилирование необходимо для транспорта большинства мРНК в цитоплазму и защищает молекулы мРНК от быстрой деградации (увеличивает время их полужизни). Лишенные поли-А участка молекулы мРНК (например, вирусные) быстро разрушаются в цитоплазме клеток эукариот рибонуклеазами.
Сплайсинг

Схема сплайсинга, в процессе которого пре-мРНК созревает в зрелую РНК. Зелёный — нетранслируемые участки (UnTranslated Regions, UTR), синий — интроны, красный — транслируемые (кодирующие белок) участки.
Сплайсинг — это процесс, в котором из пре-мРНК удаляются участки некодирующей белок последовательности, называемые интронами; последовательности, которые остаются, включают в себя кодирующие белки нуклеотиды, и называются экзонами. Иногда продукты сплайсинга пре-мРНК могут быть соединены разными способами, позволяя одному гену кодировать несколько белков. Этот процесс называется альтернативным сплайсингом. Сплайсинг обычно производится РНК-белковым комплексом, который называется сплайсосома, но некоторые молекулы мРНК также могут катализировать сплайсинг без участия белков(см. рибозимы)[2].
Транспорт
Другое различие между эукариотами и прокариотами — транспорт мРНК. Из-за того, что эукариотические транскрипция и трансляция пространственно разделены, эукариотические мРНК должны быть выведены из ядра в цитоплазму[3]. Зрелые мРНК распознаются по наличию модификаций и покидают ядро через ядерные поры, в цитоплазме мРНК образует нуклеопротеидные комплексы — информосомы, в составе которых транспортируется к рибосомам.
Трансляция
Поскольку прокариотическая мРНК не нуждается в обработке и транспортировке, трансляция рибосомой может начаться немедленно после транскрипции. Следовательно, можно сказать, что трансляция у прокариот совмещена с транскрипцией и происходит ко-транскрипционнно.
Эукариотическая мРНК должна быть обработана и доставлена из ядра в цитоплазму, и только тогда может быть транслирована рибосомой. Трансляция может происходить как на рибосомах, находящихся в цитоплазме в свободном виде, так и на рибосомах, ассоциированных со стенками эндоплазматического ретикулума. Таким образом, у эукариот трансляция не совмещена напрямую с транскрипцией.
Регуляция трансляции
Так как у прокариот транскрипция совмещена с трансляцией, прокариотическая клетка может быстро реагировать на изменения в окружающей среде путём синтеза новых белков, то есть регуляция происходит, в основном, на уровне транскрипции. У эукариот из-за необходимости в редактировании и транспорте мРНК ответ на внешние стимулы занимает больше времени. Поэтому их синтез белка интенсивно регулируется на посттранскрипционном уровне. Не всякая зрелая мРНК транслируется, поскольку в клетке существуют механизмы регуляции экспрессии белков на пост-транскрипционном уровне, например, РНК-интерференция.
Разрушение
По прошествии некоторого времени, определяемого её нуклеотидной последовательностью, в частности, длиной полиаденинового участка на 3′ конце молекулы, мРНК разрушается на составляющие её нуклеотиды с участием РНКаз. Как правило, разрушение начинается с удаления кэпа на 5′ конце, полиаденинового хвоста на 3′ конце и затем нуклеазы одновременно разрушают мРНК в 5′ ->3′ и 3′ ->5′ направлении. мРНК, в которой сигнал завершения синтеза белка, стоп-кодон, в результате ошибки транскрипции находится в середине кодирующей последовательности, подвержена особой быстрой форме деградации, НМД, (nonsense-mediated decay).
Строение зрелой мРНК

Схема строения зрелой эукариотической мРНК
Зрелая мРНК состоит из нескольких участков, различающихся по функциям: «5′ кэп», 5′ нетранслируемая область, кодирующая (транслируемая) область, 3′ нетранслируемая область и 3′ полиадениновый «хвост».
5′ Кэп
5′ кэп (или кап) (от англ. cap — шапочка) — это модифицированный гуанидиновый нуклеотид, который добавляется на 5′ (передний) конец незрелой мРНК. Эта модификация очень важна для узнавания мРНК при инициации трансляции, а также для защиты от 5’нуклеаз — ферментов, разрушающих цепи нуклеиновых кислот с незащищённым 5′-концом.
Кодирующие области
Кодирующие области состоят из кодонов — следующих непосредственно друг за другом последовательностей из трёх нуклеотидов, каждая из которых соответствует в генетическом коде определённой аминокислоте или началу и концу синтеза белка. Кодирующие области начинаются со старт-кодона и заканчиваются одним из трёх стоп-кодонов. Считывание последовательности кодонов и сборка на её основе последовательности аминокислот синтезируемой молекулы белка осуществляется рибосомами при участии транспортных РНК в процессе трансляции. В дополнение к кодированию белков, части кодирующих областей могут служить управляющими последовательностями. Например, вторичная структура РНК в некоторых случаях определяет результат трансляции.
Моноцистронная и полицистронная мРНК
мРНК называют моноцистронной, если она содержит информацию, необходимую для трансляции только одного белка (один цистрон). Полицистронная мРНК кодирует несколько белков. Гены (цистроны) в такой мРНК разделены интергенными, некодирующими последовательностями. Полицистронные мРНК характерны для прокариот и вирусов, у эукариот большая часть мРНК является моноцистронной [4]. Полицистронные мРНК встречаются у эукариот в митохондриях.
Нетранслируемые области
Нетранслируемые области — участки РНК, расположенные до старт-кодона и после стоп-кодона, которые не кодируют белок. Они называются 5′-нетранслируемая область и 3′-нетранслируемая область, соответственно. Эти области транскрибируются в составе того же самого транскрипта, что и кодирующий участок. Нетранслируемые области имеют несколько функций в жизненном цикле мРНК, включая регуляцию стабильности мРНК, локализации мРНК и эффективности трансляции. Стабильность мРНК может контролироваться 5′- и/или 3′-областью из-за различной чувствительности к ферментам, которые отвечают за деградацию РНК — РНКазам и регуляторным белкам, которые убыстряют или замедляют деградацию[5].
3′ полиадениновый хвост
Длинная (часто несколько сотен нуклеотидов) последовательность адениновых оснований, которая присутствует на 3′ «хвосте» мРНК эукариот, синтезируется ферментом полиаденилат-полимеразой. У высших эукариот поли-А-хвост добавляется к транскрибированной РНК, которая содержит специфическую последовательность, AAUAAA. Важность этой последовательности можно увидеть на примере мутации в гене человеческого 2-глобина, которая изменяет AAUAAA на AAUAAG, что приводит к недостаточному количеству глобина в организме[6].
Вторичная структура
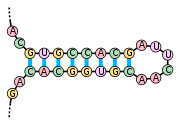
«Стебель-петля» — элемент вторичной структуры мРНК, схематично

«Псевдоузел» — элемент вторичной структуры мРНК, схематично
Кроме первичной структуры (последовательности нуклеотидов), мРНК обладает вторичной структурой. В отличие от ДНК, вторичная структура которой основана на межмолекулярных взаимодействиях (двойная спираль ДНК образована двумя линейными молекулами, соединенными друг с другом по всей длине водородными связями), вторичная структура мРНК основана на внутримолекулярных взаимодействиях (линейная молекула «складывается», и водородные связи возникают между разными участками одной и той же молекулы).
Примерами вторичной структуры могут служить стебель-петля и псевдоузел [7]
Вторичные структуры в мРНК служат для регуляции трансляции. Например, вставка в белки необычных аминокислот, селенометионина и пирролизина, зависит от стебля-петли, расположенной в 3′ нетранслируемой области. Псевдоузлы служат для программированного изменения рамки считывания генов.
В вирусных мРНК сложные вторичные структуры (Инициация трансляции»).
См. также
Ссылки
- Жизненный цикл мРНК Флэш анимация
- База данных модификаций РНК [1] (англ.)
- База данных старт- и стоп-кодонов, нетранслируемых последовательностей [2] (англ.)
- Сайт, содержащий список программ по предсказанию свойств молекул РНК [3] (англ.)
Литература
- ↑ Bruce Alberts Molecular Biology of the Cell; Fourth Edition. — New York and London: Garland Science. — ISBN ISBN 0-8153-3218-1
- ↑ Johnston W, Unrau P, Lawrence M, Glasner M, Bartel D (2001). «RNA-catalyzed RNA polymerization: accurate and general RNA-templated primer extension». Science 292 (5520): 1319-25. PMID 11358999.
- ↑ Paquin N, Chartrand P. (2008). «Local regulation of mRNA translation: new insights from the bud.». Trends Cell Biol 18: 105-11.
- ↑ Poyry, T.,Kaminski, A., Jackson R (2004). «What determines whertehr mammalian ribosomes resume scanning after translation of a short upstream open reading frame». Genes and Development 18: 62 -75.
- ↑ Kozak, M. (1983). «Comparison of initiation of protein synthesis in procaryotes, eucaryotes, and organelles.». Microbiological Reviews 47 (1): 1-45. PMID 15680349.
- ↑ Shaw, G. and Kamen, R. (1986). «A conserved AU sequence from the 3′ untranslated region of GM-CSF mRNA mediates selective mRNA degradation». Cell. 46 (5): 659-67. PMID 15680349.
- ↑ Козлов, Н. Н., Кугушев, Е. И., Сабитов, Д. И., Энеев, Т. М. «Компьютерный анализ процессов структурообразования нуклеиновых кислот».
| Типы нуклеиновых кислот | |
|---|---|
| Азотистые основания | Пурины (Аденин, Гуанин) | Пиримидины (Урацил, Тимин, Цитозин) |
| Нуклеозиды | Аденозин | Гуанозин | Уридин | Тимидин | Цитидин |
| Нуклеотиды | монофосфаты (АМФ, ГМФ, UMP, ЦМФ) | дифосфаты (АДФ, ГДФ, УДФ, ЦДФ) | трифосфаты (АТФ, ГТФ, УТФ, ЦТФ) | циклические (цАМФ, цГМФ, cADPR) |
| Рибонуклеиновые кислоты | РНК | мРНК | тРНК | рРНК | антисмысловые РНК | микроРНК | некодирующие РНК | piwi-interacting RNA | малые интерферирующие РНК | малые ядерные РНК | малые ядрышковые РНК | тмРНК |
| Дезоксирибонуклеиновые кислоты | ДНК | кДНК | Геном | msDNA | Митохондриальная ДНК |
| Аналоги нуклеиновых кислот | GNA | LNA | ПНК | TNA | Морфолино |
| Типы векторов | en:phagemid | Плазмиды | Фаг лямбда | en:cosmid | en:P1 phage | en:fosmid | BAC | YAC | HAC |
Wikimedia Foundation. 2010.
РНК-содержащие вирусы — Википедия
РНК-содержащие вирусы — это вирусы, генетический материал которых представлен РНК.[1] Обычно нуклеиновая кислота одноцепочечная (оцРНК) но может быть и двуцепочечной (дцРНК).[2] Наиболее важным заболеваниями человека, вызываемыми РНК-вирусами являются геморрагическая лихорадка Эбола, ТОРС, бешенство, простуда, грипп, гепатит C, Гепатит E, лихорадка Западного Нила, полиомиелит и корь.
Международный комитет по таксономии вирусов (ICTV) относит РНК-вирусы к группам III, IV или V по системе Классификация вирусов по Балтимору|классификации вирусов по Балтимору]] и не считает РНК-вирусами, вирусы, имеющие промежуточную стадию жизненного цикла в виде ДНК.[3] Такие вирусы, с РНК в качестве их генетического материала и имеющие промежуточную стадию жизненного цикла в виде ДНК в цикле репликации, названы ретровирусами, и выделены в группу VI классификация вирусов по Балтимору. Важные ретровирусы человека включают Вирус иммунодефицита человека#ВИЧ-1 and Вирус иммунодефицита человека#ВИЧ-2, вызывающих ВИЧ-инфекцию.
Другим термином явно исключающий группу ретровирусов из РНК-вирусов является понятие рибовирусы.[4]
Одноцепочечные РНК-вирусы и смысловая РНК[править | править код]
РНК-вирусы могут быть дополнительно классифицированы в соответствии с полярностью их РНК на одноцепочечные РНК-вирусы с отрицательной цепью[en] и одноцепочечные РНК-вирусы с положительной цепью[en], или РНК обеих типов[en] двухцепочечные РНК-вирусы[en]. Вирусы, содержащие одноцепочечную (+)РНК похожую на мРНК и таким образом может непосредственно транслирована зараженной клеткой. Поэтому (+)РНК также называется смысловой. Антисмысловая (-)РНК вируса комплиментарна мРНК и таким образом может быть превращена в (+)РНК действием РНК-зависимой РНК-полимеразой перед трансляцией. Очищенная РНК (+)РНК-содержащих вирусов сама по себе может напрямую вызывать инфекцию, хотя ее инфекционность ниже, чем у полноценных вирусных частиц. Очищенная РНК (-)РНК-содержащих вирусов неинфекционна сама по себе, так как она должна транскрибироваться в (+)РНК; каждый вирион после транскрипции может быть источником нескольких смысловых цепей (+)РНК. Двухцепочечные(±)РНК вирусы напоминают (-)РНК-содержащие вирусы тем, что они также транслируют гены с отрицательной (некодирующей) цепи РНК.[5]
Двуцепочечные РНК вирусы[править | править код]
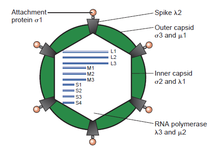 Структура вириона реовируса
Структура вириона реовирусаДвуцепочечные РНК вирусы представляет разнообразную группу вирусов отличающихся по заражаемому кругу хозяев (людей, животных, растений, грибов, и бактерий), числу фрагментов генома (от 1 до 12 сегментов), и организацией вириона (формой капсида, количеством слоев капсида, выросты, шипы, и т.п.). Члены этой группы включают ротавирусы, всемирно известные как наиболее распространенная причина детских гастроэнтеритов, и пикорнавирусы, известный во всем мире как наиболее часто встречающийся вирус в фекалиях людей и животных с признаками диареи или без них. Вирус болезни синего языка[en] является экономически важным возбудителем болезни коз и овец. В прошлые годы, достигнут большой прогресс в определении атомной и субнанометровой структуры ряда ключевых вирусных белков и капсидов вирионов нескольких дцРНК-вирусов, подчеркивая существенные параллели в структуре и механизмах процессов репликации многих из этих вирусов.[2]
Частота мутаций[править | править код]
РНК вирусы имеют очень высокую частоту мутаций по-сравнению с ДНК-вирусами,[6] потому что вирусная РНК-зависимая РНК-полимераза не имеет способности (3′-5′-экзонуклеазной гидролитической активности) к коррекции ошибок, как ДНК-полимеразы.[7] Это одна из причин того, почему так трудно сделать эффективную вакцину для предотвращения болезней, вызываемых РНК-вирусами.[8] Ретровирусы также имеют высокую частоту мутаций, даже несмотря на то, что их промежуточная форма генома в виде ДНК интегрируется в геном хозяина (и, следовательно, после интеграции в геном, во время репликации, подвергается корректирующей проверке ДНК-полимеразой хозяина). Высокая частота мутаций обусловлена высокой частотой ошибок во время обратной транскрипции, до стадии интеграции в геном хозяина. Ошибки при этом встраиваются в обе цепи ДНК.[9] Некоторые гены РНК-вирусов важны для их репликации и мутации в них не допустимы. Например, участок генома вируса гепатита С кодирующий коровый белок, высоко консервативен,[10] потому что он содержит структурную РНК, вовлеченную в образование участка внутренней посадки рибосомы.[11]
РНК-вирусы животных классифицированы по ICTV. Выделяют три различные группы РНК-вирусов в зависимотси от структуры их генома и метода репликации:
- Двуцепочечные РНК-вирусы (группа III) содержат от одной до нескольких разных молекул РНК каждая из которых кодирует один или несколько вирусных белков.
- Одноцепочечные (+)РНК-вирусы (группа IV) используют свой геном напрямую как мРНК, при трансляции на рибосомах хозяина в один белок, который модифицируется белками хозяина и самого вируса для образования различных белков необходимых для процесса репликации. Один из них это РНК-зависимая РНК полимераза (РНК репликаза), которая копирует РНК вируса переводя ее в двуцепочечную форму, способную к репликации. В свою очередь дцРНК служит матрицей для синтеза новых вирусных РНК-геномов.
- Одноцепочечные (-)РНК-вирусы (группа V) должны обеспечить копирование их генома РНК репликазой для формирования смысловой (+)нити РНК. Это означает, что вирус уже должен содержать готовый формент РНк репликазу вместе со своим геномом. Смысловая (+) нить РНК затем выступает в качестве мРНК, которая транслируется в белки рибосомами хозяина.
Ретровирусы (группа VI) имеют одноцепочечный РНК-геном но, в общем, не относятся к РНК-вирусам потому что они используют промежуточную стадию в виде ДНК молекулы для репликации. Обратная транскриптаза, вирусный белок поставляемый самим вирусом, после распаковки вирусной частицы, конвертирует вирусную РНК в комплементарную цепь ДНК, которая после синтеза второй ДНК цепи превращается в двуцепочечную молекулу вирусной ДНК. После этого ДНК интегрируется в геном хозяина используя вирусный белок интегразу, экспрессия закодированных генов может приводить к формированию новых вирионов.
Отчасти это связано с высокой частотой мутаций, которым подвергаются эти геномы. Классификация основана главным образом на типе генома (двухцепочечный, отрицательный или положительный одноцепочечный), а также количестве и организации генов. В настоящее время известно 5 порядков и 47 семейств РНК-вирусов. Есть также много неклассифицированных видов и родов. Родственные но отличающиеся от РНк-вирусов вироиды и РНК вирусы-сателлиты. Они не относятся к РНК-вирусам и описаны отдельно.
Изучение нескольких тысяч РНК-вирусов показало существование по крайней мере пяти главных таксонов: »Levivirus» и родственные группы; супергруппа »Picornavirus»; супергруппа »Alphavirus» вместе с супергруппой »Flavivirus»; дцРНК вирусы; и оц(-)РНК вирусы.[12] Группа лентивирусов является предковой для всех оставшихся групп РНК-вирусов. Следующий большой водораздел пролегает между группой пикорнавирусов и остальными вирусами. Вирусы с дцРНК геномом похоже эволюционировали от оц(+)РНК вирусного предка, а оц(-)РНК-вирусы возникли внутри дцРНК-вирусов. Наиболее близкородственные (-)РНК вирусам: реовирусы.
Одноцепочечные РНК-вирусы с положительной цепью[править | править код]
Это самая большая группа РНК-вирусов[13] с 30 семействами. Предпринималась попытка сгруппировать их в более крупные порядки на основе анализа их РНК-зависимой РНК-полимеразы. На сегодняшний момент это предложение не получило широкого распространения из-за сомнений в пригодности использования одного гена для классификации клады. В предложенной классификации выделены три группы[14]:
- Пикорнаподобная группа (Picornavirata): Bymoviruses, Comoviruses, Nepoviruses, Nodaviruses, Picornaviruses, Potyviruses, Sobemoviruses и подмножество Luteoviruses (beet western yellows virus и potato leafroll virus).
- Группа, похожая на флавивирусы (Flavivirata): Carmoviruses, Dianthoviruses, Flaviviruses, Pestiviruses, Statoviruses, Tombusviruses, одноцепочечные РНК-бактериофаги, вирус гепатита С и подмножество лютеовирусов (вирус желтой карликовости ячменя).
- Группа, похожая на альфавирусы (Rubivirata): Alphaviruses, Carlaviruses, Furoviruses, Hordeiviruses, Potexviruses, Rubiviruses, Tobraviruses, Tricornaviruses, Tymoviruses, apple chlorotic leaf spot virus, beet yellows virus и вирус гепатита Е.
Было предложено разделение альфа-подобной (Sindbis-подобной) супергруппы на основе нового домена, расположенного вблизи N-концов белков, участвующих в репликации вируса. [15]. Предлагаемые две группы:
- Группа ‘Альтовирус’: Alphaviruses, Furoviruses, вирус гепатита Е, Hordeiviruses, Tobamoviruses, Tobraviruses, Tricornaviruses и, возможно, Rubiviruses;
- Группа ‘Типовирус’: apple chlorotic leaf spot virus, Carlaviruses, Potexviruses и Tymoviruses
В будущем альфа-подобную (Sindbis-подобную) супергруппу можно будет разделить на три группы: Rubi-подобные, Tobamo-подобные, и Tymo-подобные вирусы.[16]
Дополнительная работа позволила выявить пять групп РНК-вирусов с положительной цепью, содержащих 14 порядков с 31 семейством и 48 родов (из них 17 семейств с 30 родами относятся к вирусам растений).[17]. Этот анализ предполагает, что альфавирусы и флавивирусы могут быть разделены на два семейства — Togaviridae и Flaviridae, соответственно, — но выделение других таксономических единиц, таких как Pestiviruses, вирус гепатита С, Rubiviruses, вирус гепатита Е и Arteriviruses, могут быть неверным. Коронавирусы и торовирусы, по-видимому, представляют собой отдельные семейства в разных порядках, а не отдельные роды того же семейства, как они классифицированы в настоящее время. Лютеовирусы, по-видимому, представляют собой два семейства, а не одно, а apple chlorotic leaf spot virus, по-видимому, является не Closterovirus, а новым родом Potexviridae.
- Эволюция
Эволюция пикорнавирусов, основанная на анализе их РНК-полимераз и геликаз, по-видимому, на сегодняшний день ведет свое происхождение от супергруппы эукариот.[18]. Их предполагаемые предки включают бактериальные ретроэлементы группы II, семейство протеаз HtrA и ДНК-бактериофаги.
Partitiviruses связаны и, возможно, произошли от предка тотивируса.[19]
Гиповирусы и барнавирусы, по-видимому, имеют общую родословную с линиями потивирусов и собемовирусов соответственно.[19]
Двуцепочечные РНК-вирусы[править | править код]
Анализ также предполагает, что дцРНК-вирусы не связаны друг с другом, а вместо этого принадлежат к четырем дополнительным классам — Birnaviridae, Cystoviridae, Partitiviridae и Reoviridae — и одному дополнительному отряду (Totiviridae), одного из классов оц(+)РНК-вирусов и относятся к той же группе , что и РНК-вирусы с положительной цепью.
Одно исследование показало, что существует две большие клады: одна включает семейства Caliciviridae, Flaviviridae и Picornaviridae, а вторая — семейства Alphatetraviridae, Birnaviridae и Cystoviridae, Nodaviridae и Permutotretraviridae.[20]
Одноцепочечные РНК-вирусы с негативной цепью[править | править код]
Эти вирусы имеют сегментированный геном, начиная от одного до восьми сегментов РНК. Несмотря на их разнообразие, они, возможно, возникли у предка членистоногих и дивергировали вместе с ним.[21]
Вирусы-сателлиты[править | править код]
Также известен ряд вирусов-сателлитов — вирусов, для завершения жизненного цикла которых требуется помощь другого вируса. Их таксономия еще не согласована. Следующие четыре рода были предложены для сателлитных одноцепочечных РНК-вирусов со смысловой цепью, которые заражают растения: Albetovirus, Aumaivirus, Papanivirus и Virtovirus[22]. Семейство Sarthroviridae, включающее род Macronovirus — были предложены для сателлитных одноцепочечных РНК-вирусов со смысловой цепью, которые заражают членистоногих.
В этой группе насчитывается двенадцать семейств, а также ряд неклассифицированных родов и видов.[7]
В этой группе есть три порядка и 34 семейства. Кроме того, существует ряд неклассифицированных и родов.
Вирусы-сателлиты
Также был описан неклассифицированный астровирусный / гепевирусоподобный вирус.[24]
За исключением гепатита D, эта группа вирусов была помещена в один тип — Negarnaviricota. Этот тип был разделен на два подтипа — Haploviricotina и Polyploviricotina. Внутри подтипа Haploviricotina в настоящее время признаются четыре класса: Chunqiuviricetes, Milneviricetes, Monjiviricetes и Yunchangviricetes. В подтипе Polyploviricotina выделяются два класса: Ellioviricetes и Insthoviricetes.
В настоящее время в этой группе признано 6 классов, 7 порядков и 24 семейства. Чаксть вилов и родов еще не классифицированы.[7]
- Неклассифицированные роды:
Большинство вирусов, поражающих грибы, представляют собой двухцепочечные РНК-вирусы. Описано небольшое количество одноцепочечных РНК-вирусов с положительной цепью. В одном сообщении высказывается предположение о возможности заражения одноцепочечным вирусом с негативной цепью.[27]
- ↑ MeSH, retrieved on 12 April 2008.
- ↑ 1 2 Patton JT (editor). Segmented Double-stranded RNA Viruses: Structure and Molecular Biology (англ.). — Caister Academic Press (англ.)русск., 2008. — ISBN 978-1-904455-21-9.
- ↑ Listing in Taxonomic Order—Index to ICTV Species Lists (неопр.). Дата обращения 11 апреля 2008.
- ↑ Drake J. W., Holland J. J. Mutation rates among RNA viruses (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1999. — November (vol. 96, no. 24). — P. 13910—13913. — doi:10.1073/pnas.96.24.13910. — Bibcode: 1999PNAS…9613910D. — PMID 10570172.
- ↑ Nguyen M., Haenni A. L. Expression strategies of ambisense viruses (неопр.) // Virus Res.. — 2003. — Т. 93, № 2. — С. 141—150. — doi:10.1016/S0168-1702(03)00094-7. — PMID 12782362.
- ↑ Sanjuan, R.; Nebot, M. R.; Chirico, N.; Mansky, L. M.; Belshaw, R. Viral Mutation Rates (англ.) // Journal of Virology (англ.)русск. : journal. — 2010. — Vol. 84, no. 19. — P. 9733—9748. — ISSN 0022-538X. — doi:10.1128/JVI.00694-10. — PMID 20660197.
- ↑ 1 2 3 Klein, Donald W.; Prescott, Lansing M.; Harley, John. Microbiology (неопр.). — Dubuque, Iowa: Wm. C. Brown, 1993. — ISBN 978-0-697-01372-9.
- ↑ Steinhauer D. A., Holland J. J. Rapid evolution of RNA viruses (англ.) // Annu. Rev. Microbiol.. — 1987. — Vol. 41. — P. 409—433. — doi:10.1146/annurev.mi.41.100187.002205. — PMID 3318675.
- ↑ Boutwell C. L., Rolland M. M., Herbeck J. T., Mullins J. I., Allen T. M. Viral Evolution and Escape during Acute HIV-1 Infection (англ.) // J. Infect. Dis. : journal. — 2010. — October (vol. 202, no. Suppl 2). — P. S309—14. — doi:10.1086/655653. — PMID 20846038.
- ↑ Bukh J., Purcell R. H., Miller R. H. Sequence analysis of the core gene of 14 hepatitis C virus genotypes (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1994. — August (vol. 91, no. 17). — P. 8239—8243. — doi:10.1073/pnas.91.17.8239. — Bibcode: 1994PNAS…91.8239B. — PMID 8058787.
- ↑ Tuplin A., Evans D. J., Simmonds P. Detailed mapping of RNA secondary structures in core and NS5B-encoding region sequences of hepatitis C virus by RNase cleavage and novel bioinformatic prediction methods (англ.) // Journal of General Virology (англ.)русск. : journal. — Microbiology Society (англ.)русск., 2004. — October (vol. 85, no. Pt 10). — P. 3037—3047. — doi:10.1099/vir.0.80141-0. — PMID 15448367.
- ↑ Wolf YI, Kazlauskas D, Iranzo J, Lucía-Sanz A, Kuhn JH, Krupovic M, Dolja VV, Koonin EV (2018) Origins and Evolution of the Global RNA Virome. MBio 9(6) pii: e02329-18
- ↑ Francki, R.I.B; Fauquet, C. M.; Knudson, D. L.; Brown, F. Classification and nomenclature of viruses. Fifth report of the International Committee on Taxonomy of Viruses, Archives of Virology (Suppl. 2) (англ.). — 1991. — ISBN 978-3-7091-9163-7.
- ↑ Koonin E. V. The phylogeny of RNA-dependent RNA polymerases of positive-strand RNA viruses (англ.) // Journal of General Virology (англ.)русск. : journal. — Microbiology Society (англ.)русск., 1991. — September (vol. 72, no. Pt 9). — P. 2197—2206. — doi:10.1099/0022-1317-72-9-2197. — PMID 1895057.
- ↑ Rozanov M. N., Koonin E. V., Gorbalenya A. E. Conservation of the putative methyltransferase domain: a hallmark of the ‘Sindbis-like’ supergroup of positive-strand RNA viruses (англ.) // Journal of General Virology (англ.)русск. : journal. — Microbiology Society (англ.)русск., 1992. — August (vol. 73, no. Pt 8). — P. 2129—2134. — doi:10.1099/0022-1317-73-8-2129. — PMID 1645151.
- ↑ Koonin E. V., Dolja V. V. Evolution and taxonomy of positive-strand RNA viruses: implications of comparative analysis of amino acid sequences (англ.) // Crit. Rev. Biochem. Mol. Biol. (англ.)русск. : journal. — 1993. — Vol. 28, no. 5. — P. 375—430. — doi:10.3109/10409239309078440. — PMID 8269709.
- ↑ Ward C. W. Progress towards a higher taxonomy of viruses (неопр.) // Res Virol. — 1993. — Т. 144, № 6. — С. 419—453. — doi:10.1016/S0923-2516(06)80059-2. — PMID 8140287.
- ↑ Koonin E. V., Wolf Y. I., Nagasaki K., Dolja V. V. The Big Bang of picorna-like virus evolution antedates the radiation of eukaryotic supergroups (англ.) // Nat. Rev. Microbiol. : journal. — 2008. — December (vol. 6, no. 12). — P. 925—939. — doi:10.1038/nrmicro2030. — PMID 18997823.
- ↑ 1 2 Ghabrial SA (1998) Origin, adaptation and evolutionary pathways of fungal viruses. Virus Genes 16(1):119-131
- ↑ Gibrat J. F., Mariadassou M., Boudinot P., Delmas B. Analyses of the radiation of birnaviruses from diverse host phyla and of their evolutionary affinities with other double-stranded RNA and positive strand RNA viruses using robust structure-based multiple sequence alignments and advanced phylogenetic methods (англ.) // BioMed Central (англ.)русск. : journal. — 2013. — Vol. 13. — P. 154. — doi:10.1186/1471-2148-13-154. — PMID 23865988.
- ↑ Li CX, Shi M, Tian JH, Lin XD, Kang YJ, Chen LJ, Qin XC, Xu J, Holmes EC, Zhang YZ (2015) Unprecedented genomic diversity of RNA viruses in arthropods reveals the ancestry of negative-sense RNA viruses. Elife 4. doi: 10.7554/eLife.05378.
- ↑ Krupovic M., Kuhn J. H., Fischer M. G. (2016). A classification system for virophages and satellite viruses. Arch Virol 161(1): 233—247 doi:10.1007/s00705-015-2622-9.
- ↑ Adams M. J., Antoniw J. F., Kreuze J. Virgaviridae: a new family of rod-shaped plant viruses (англ.) // Arch Virol (англ.)русск. : journal. — 2009. — Vol. 154, no. 12. — P. 1967—1972. — doi:10.1007/s00705-009-0506-6. — PMID 19862474.
- ↑ Pankovics P, Boros Á, Kiss T, Engelmann P, Reuter G (2019) Genetically highly divergent RNA virus with astrovirus-like (5′-end) and hepevirus-like (3′-end) genome organization in carnivorous birds, European roller (Coracias garrulus). Infect Genet Evol
- ↑ Virus Taxonomy: 2018 Release (неопр.). International Committee on Taxonomy of Viruses. Дата обращения 13 ноября 2018.
- ↑ Mihindukulasuriya, K. A.; Nguyen, N. L.; Wu, G.; Huang, H. V.; Travassos da Rosa, A. P.; Popov, V. L.; Tesh, R. B.; Wang, D. Nyamanini and Midway viruses define a novel taxon of RNA viruses in the Порядок Mononegavirales (англ.) // J. Virol. (англ.)русск. : journal. — 2009. — Vol. 83, no. 10. — P. 5109—5116. — doi:10.1128/JVI.02667-08. — PMID 19279111.
- ↑ Kondo, H.; Chiba, S.; Toyoda, K.; Suzuki, N. Evidence for negative-strand RNA virus infection in fungi (англ.) // Virology : journal. — 2012. — Vol. 435, no. 2. — P. 201—209. — doi:10.1016/j.virol.2012.10.002. — PMID 23099204.