понятие, механизм. Гетеро- и гомометрические механизмы регуляции сократимости.
Сердечная мышца реагирует на раздражители нарастающей силы по закону «все или ничего». Это обусловлено ее морфологическими особенностями. Между отдельными мышечными клетками сердечной мышцы имеются так называемые вставочные диски, или участки плотных контактов — нексусы, образованные участками плазматических мембран двух соседних миокардиальных клеток. В некоторых участках плазматические мембраны, образующие контакт, прилегают друг к другу так близко, что кажутся слившимися. Мембраны на уровне вставочных дисков обладают очень низким электрическим сопротивлением и поэтому возбуждение распространяется от волокна к волокну беспрепятственно, охватывая миокард целиком. Поэтому сердечную мышцу, состоящую из морфологически разъединенных, но функционально объединенных мышечных волокон, принято считать функциональным синцитием.
Способность сердечной мышцы сокращаться только по тину одиночного сокращения обеспечивает выполнение сердцем основной гемодинамической функции — насоса. Сокращения сердца по типу тетануса делали бы невозможным ритмическое нагнетание крови в кровеносные сосуды. Именно это и происходит при фибрилляции волокон миокарда и мерцательной аритмии сердца.
Серию последовательных явлений в клетке миокарда, начинающихся с пускового механизма сокращения — потенциала действия (ПД) и завершающихся укорочением миофибрилл, называют сопряжением возбуждения и сокращения. При распространении ПД по мембране ионы кальция поступают к сократительным белкам, в основном, из межклеточного пространства и вызывают те же процессы взаимодействия актиновых и миозиновых протофибрилл, что и в скелетном мышечном волокне. Расслабление кардиомиоцита также обусловлено удалением кальция кальциевым насосом из протофибриллярного пространства в межклеточную среду.
Важным процессом в сокращении кардиомиоцита является вход ионов кальция в клетку во время ПД. Наряду с тем, что входящий в клетку кальций увеличивает длительность ПД и, как следствие, продолжительность рефракторного периода, он является важнейшим фактором в регуляции силы сокращения сердечной мышцы.
Так, удаление ионов кальция из межклеточных пространств приводит к полному разобщению процессов возбуждения и сокращения — потенциал действия остается практически в неизменном виде, а сокращения кардиомиоцита не происходит.
Миогенная авторегуляция включает в себя гетерометрический и гомеометрический механизмы. Гетерометрический механизм опосредован внутриклеточными взаимодействиями и связан с изменением взаиморасположения актиновых и миозиновых нитей в миофибриллах кардиомиоцитов при растяжении миокарда кровью, поступающей в полости сердца. Растяжение миокардиоцитов приводит к увеличению количества миозиновых мостиков, способных соединить миозиновые и актиновые нити во время сокращения. Чем более растянут кардиомиоцит, тем на большую величину он может укоротиться при сокращении, и тем более сильным будет это сокращение. Этот вид регуляции был установлен на сердечно-легочном препарате и сформулирован в виде «закона сердца» или закона Франка-Старлинга. Согласно этому, закону, чем больше миокард растянут во время диастолы, тем больше сила последующего сокращения (систолы). Предсистолическое растяжение миокарда обеспечивается дополнительным объемом крови, нагнетаемым в желудочки во время систолы предсердии. При утомлении сердечной мышцы и длительной нагрузки (например, при гипертонии) этот закон проявляется только в том случае, если сердечная мышца растягивается значительно больше, чем обычно. Однако, величина минутного объема сердца и в этих состояниях длительное время удерживается на нормальном уровне. При дальнейшем нарастании утомления или нагрузки этот показатель уменьшается.
Гомеометрическая авторегуляция сердца связана с определенными межклеточными отношениями и не зависит от пред систолического его растяжения. Большую роль в гомеометрической регуляции играют вставочные диски — нексусы, через которые миокардиоциты обмениваются ионами и информауией. Реализуется данная форма регуляции в виде «эффекта Анрепа» — увеличение силы сердечного сокращения при возрастании сопротивления в магистральных сосудах.
Другим проявлением гомеометрической регуляции является так называемая ритмоинотропная зависимость: изменение силы сердечных сокращений при изменении частоты. Это явление обусловлено изменением длительности потенциала действия миокардиоцитов и, следовательно, изменением количества экстрацеллюлярного кальция, входящего в миокардиоцит при развитии возбуждения.
3. Задача. Если при ярком освещении пристально посмотреть на какой-либо предмет, а потом закрыть глаза, то на протяжении некоторого времени можно видеть этот предмет. Каким свойством нервных центров объясняется это явление?
Последействие, это запаздывание окончания рефлекторного ответа после прекращения действия раздражителя. Связано с циркуляцией нервных импульсов по замкнутым цепям нейронов.
Посттетаническая потенциация, это усиление рефлекторной реакции в результате длительного возбуждения нейронов центра. Под влиянием многих серий нервных импульсов, проходящих с большой частотой через синапсы, выделяется большое количество нейромедиатора в межнейронных синапсах. Это приводит к прогрессирующему нарастанию амплитуды возбуждающего постсинаптического потенциала и длительному (несколько часов) возбуждению нейронов.
149Механизм работы сердца — SportWiki энциклопедия
В отличие от скелетных мышц, невозможно вызвать произвольное сокращение сердечной мускулатуры. Одной из важнейших характеристик сердечных мышц является так называемый автоматизм сокращений. Именно поэтому сердце продолжает сокращаться вне организма человека, т. к. возбуждающие потенциалы формируются в самом сердце. Первичным центром формирования потенциалов действия является синусовый (синоатриальный) узел. В норме в нем возникает возбуждение с частотой примерно 60-90 импульсов в минуту. Из синусового узла возбуждение распространяется радиально через предсердия в атриовентрикулярный узел (АВ-узел), где проведение возбуждения несколько замедляется. Затем возбуждение достигает пучка Гиса, где по ножкам переходит в желудочки в волокна Пуркинье. При нарушении работы синусового узла его функцию берет на себя АВ-узел, генерирующий импульсы с частотой 40-60 в минуту (Maurer, 2006; Рис. 1).
Рис. 1. Строение проводящей системы сердцаЭлектрокардиограмма[править | править код]
Формирование возбуждения, его распространение и затухание в различных областях сердца происходит при разности потенциалов в возбужденных и невозбужденных клетках, равной 120 мВ. Эта разность потенциалов может быть зарегистрирована на поверхности тела с помощью электродов, установленных на определенные точки отведения, что позволяет получить картину распространения возбуждения в сердце.
Отведения электрокардиограммы (ЭКГ) имеют определенную форму, которая связана с функцией сердца — выделяют постоянные волны, интервалы и зубцы. Расстояние между двумя соседними зубцами R называется сердечным циклом (Рис. 2).
Рис. 2. Электрокардиограмма сердцаЧастота сердечных сокращений[править | править код]
В ответ на 60-90 потенциалов действия, возникающие в покое в синусовом узле, в миокарде происходят мышечные сокращения с различной частотой в минуту. При снижении частоты сердечных сокращений ниже 60 в минуту говорят о брадикардии (De Marees, 2003), а при повышении выше 100 в минуту, например, при физической нагрузке, — о (нагрузочной) тахикардии.
Нервы сердца[править | править код]
Автономная работа сердца регулируется симпатическими и парасимпатическими волокнами (вегетативной нервной системы), следующими от центров сердечно-сосудистой деятельности через спинной мозг к сердцу. За счет возбуждающего действия симпатической системы и тормозящего действия парасимпатической системы регулируется работа сердца в зависимости от потребностей организма в кислороде. В нервных окончаниях высвобождаются вещества — переносчики сигнала, нейротрансмиттеры, которые стимулируют передачу сигнала на клетки сердечных мышц. К нейротрансмиттерам относят норадреналин и в меньшей степени адреналин для симпатической нервной системы и ацетилхолин для парасимпатической. Механизм их действия заключается в связывании со специфическими адренергическими или холинергическими рецепторами.
Запомните:
Вегетативная нервная система | Нейромедиатор | Эффекты |
Симпатическая | Норадреналин, адреналин | Хронотропный (частота), ионотропный (сила) и дромотропный (проводимость) |
Парасимпатическая | Ацетилхолин |
Нервы сердца не могут вызывать изолированные сердечные сокращения из-за механизмов автоматии, однако оказывают на функцию сердца различные влияния. Выделяют три вида влияний на сердце:
- хромотропное действие — влияние на частоту сердечных сокращений;
- ионотропное действие — влияние на силу сердечных сокращений;
- дромотропное действие — влияние на скорость проведения возбуждения (проводимость).
Механизмом симпатической регуляции частоты сердечных сокращений является повышение частоты генерации импульсов в синусовом узле и ускорение проведения возбуждения в АВ-узле. Кроме этого, при активации симпатической системы повышается сила сердечных сокращений. Отрицательный ионотропный эффект парасимпатической системы выражен очень мало, т. к. она иннервирует только предсердия и не влияет на желудочки. Частота сердечных сокращений при отсутствии влияний нервной системы составляет 100-120 ударов в минуту, а фактическая меньшая частота в покое объясняется преобладанием парасимпатических влияний на синусовый узел. Активность симпатической и парасимпатической системы также называют тонусом. Если после периода отдыха при физической нагрузке необходимо быстрое повышение частоты сокращений, вначале происходит уменьшение парасимпатического тонуса. Для дальнейшего увеличения частоты (> 120 ударов в минуту) и ударного объема необходимо повышение симпатического тонуса.
Запомните: Парасимпатическая система иннервирует только предсердия и не влияет на желудочки.
Электромеханическое сопряжение[править | править код]
Рис. 3. Потенциалы действия сердца и скелетных мышцПотенциалы действия сердечных мышц в покое отличаются значительно большей длительностью, чем потенциалы действия скелетной мускулатуры — 200 мс по сравнению с 1-2 мс. Это объясняется тем, что, в отличие от нервов или скелетных мышц, при возникновении потенциала действия в сердце помимо быстрого тока через Nа+-каналы открываются медленные Са2+-каналы, которые с большой задержкой прекращают ток ионов. В результате этого возникают длинная фаза плато (Рис. 3) и удлиненная фаза рефрактерности (время, в течение которого невозможно новое возбуждение). Удлинение потенциала действия играет роль защиты от так называемых циркулирующих волн возбуждения и тетанических сокращений (постоянное сокращение).
В отличие от скелетных мышц, в сердце невозможно суммирование мышечных сокращений (суперпозиция). Сердце представляет собой одну большую моторную единицу, поэтому невозможно повышение силы сокращений за счет привлечения других моторных единиц, находящихся в состоянии покоя, как в скелетных мышцах. Тем не менее сокращения сердца все же подобны работе скелетной мускулатуры. Ключевую роль в электромеханическом сопряжении играют ионы Са2+. Входной поток ионов Са2+ или норадренергическая стимуляция через β-адренорецепторы приводят к высвобождению ионов Са2+ из продольных трубочек саркоплазматического ретикулума, которые присоединяются к тропонину С и приводят к образованию мостовидных связей между актином и миозином (Maurer, 2006). После сокращения ионы Са2+ вновь выводятся во внеклеточное пространство Nа+-Са2+-насосом или специфическими Са2+ -насосами (SERCA — саркоэндоплазматический ретикулум Са-АТФаза) в саркоплазматический ретикулум. Продолжительность сокращения зависит от длительности потенциала действия, а сила сокращений — от концентрации Са2+ в цитоплазме во время этой фазы. Таким образом, вещества, влияющие на входной поток Са2+, могут изменять силу сердечных сокращений. К таким веществам, например, относится адреналин, который увеличивает поток ионов Са2+ в цитоплазму (Tortora, Derrickson, 2006).
Запомните: Под периодом рефрактерности понимают время, в течение которого невозможно новое сокращение. Рефрактерность является защитным механизмом против тетанических сокращений для обеспечения насосной функции сердца. Ключевую роль в электромеханическом сопряжении играют ионы Са2+.
Этим объясняется механизм регуляции сердечной деятельности симпатической нервной системой. Механизм парасимпатической регуляции заключается в выведении К+ из клетки, что приводит к накоплению отрицательного заряда с внутренней стороны клеточной мембраны и затруднению деполяризации и формирования потенциала действия.
Ударный объем[править | править код]
Объем крови (в миллилитрах), выбрасываемый во время одной систолы из левого желудочка, называется ударным объемом. В покое он составляет приблизительно 70-100 мл и сильно зависит от положения тела. В положении лежа улучшается диастолическое наполнение сердца, в то время как повышение ударного объема в ответ на физическую нагрузку оказывает меньшее влияние. Ударный объем зависит от следующих факторов:
1) преднагрузки — давления наполнения в конце диастолы;
2) постнагрузки — давления в сосудах, отходящих от сердца;
3) силы сокращений сердца.
Сердечный цикл[править | править код]
Сердце работает по принципу помпы. Выделяют две фазы функции сердца — фазу сокращения (систола) и фазу расслабления (диастола). Диастола в покое практически в 2 раза длиннее систолы, а при физической нагрузке это соотношение становится обратным. В систоле выделяют фазы напряжения (около 50 мс) и изгнания (210 мс), а в диастоле — фазы расслабления (60 мс) и наполнения (500 мс). Для поддержания тока крови из вен в предсердия, желудочки и затем в артерии предсердия и желудочки сокращаются попеременно. Таким образом, систола предсердий происходит во время диастолы желудочков и заканчивается перед их систолой. При этом клапаны сердца закрываются и предотвращают обратный ток крови. Открытие и закрытие клапанов обусловлено их анатомическим строением и разницей давлений на обеих сторонах клапана. Во время фазы напряжения давление в желудочках повышается, что приводит к закрытию створчатых клапанов. При этом полулунные клапаны остаются закрытыми, т. к. объем желудочка не изменяется, однако, если давление в левом желудочке превышает давление в аорте, полулунный клапан аорты пассивно открывается. В этот момент начинается фаза изгнания, при которой объем желудочка уменьшается на ударный объем. В эту фазу артериальное давление повышается до максимального систолического давления (справа: около 20-25 мм рт. ст.; слева: около 120 мм рт. ст.). Около 60 % крови из конечного диастолического объема в покое выбрасывается как ударный объем, а остаток называется остаточным объемом. Доля ударного объема в процентном соотношении от конечного диастолического объема называется фракцией выброса. В начале диастолы все клапаны закрыты, благодаря чему повышается внутрижелудочковое давление. По этой причине фаза расслабления в начале диастолы называется изоволюметрической, т. е. объем камер сердца не изменяется. Когда давление в желудочках превышает давление в предсердиях, открываются створчатые клапаны и происходит опорожнение желудочков. В конце диастолы желудочков происходит сокращение предсердий. Наполнение желудочков происходит преимущественно за счет механизма сдвига клапанов. Во время систолы желудочков клапаны сердца сдвигаются к верхушке сердца, при этом кровь засасывается в предсердия. Во время диастолы желудочков клапаны вновь смещаются к предсердиям, при этом клапаны открываются и кровь поступает в желудочки. Фазы, при которых не происходит изменения объема камер, протекают относительно быстро (50-60 мс), а фазы с изменением давления и объема — значительно медленнее.
Механизм Франка—Старлинга[править | править код]
При изменении венозного оттока к правому отделу сердца вследствие различных факторов (например, изменения положения тела) благодаря механизму Франка—Старлинга происходит изменение ударного объема для сохранения постоянства тока крови в малом и большом кругах кровообращения. При этом выравнивается разница в ударных объемах между правой и левой половинами сердца. Этот механизм описывает зависимость силы сердечных сокращений от растяжения сердечной стенки. Повышение ударного объема происходит без участия вегетативной нервной системы (экстракардиальная система), поэтому он называется интракардиальным адаптационным механизмом.
Механизм мышечного сокращения
Удалось установить, что при мышечном сокращении:
длина протофибрилл не меняется,
диски А также не меняются (не укорачиваются),
укорачиваются диски I,
укорачиваются или исчезают зоны Н.
Укорочение дисков I и зон Н зависит от того, что при мышечном сокращении концы тонких протофибрилл в зонах перекрытия глубже выдвигаются в промежутки между толстыми, занимая зоны Н. При этом белок актин тонких протофибрилл связывается с белком миозином толстых, образуя новый актомиозиновый белковый комплекс. В момент расслабления мышцы этот комплекс распадается на свои составные части – белки актин и миозин.
На основании того, что при мышечном сокращении толстые и тонкие протофибриллы скользят друг относительно друга, теория мышечного сокращения получила название: теория скользящих нитей.
Т – система – это выпячивания сарколеммы внутрь волокна. Они охватывают каждую миофибриллу на границе А и I дисков. Эта система обеспечивает передачу нервных импульсов ко всем миофибриллам одновременно.
Источником регенерации поперечнополосатых мышечных волокон являются малодифференцированные клетки (клетки – сателлиты), которые располагаются в составе сарколеммы между плазмалеммой и базальной мембранной. Эти клетки превращаются в миобласты, которые сливаясь друг с другом образуют мышечное волокно. Между отдельными мышечными волокнами располагается сеть коллагеновых, эластических и ретикулярных волокон, а также некоторые соединительнотканные клетки – это эндомизий.
Между отдельными пучками мышечных волокон располагаются прослойки соединительной ткани – это перимизий. На периферии мышц – толстый слой соединительной ткани – эпимизий.
Сердечная мышечная ткань
Сердечные мышечные волокна состоят из продольно ориентированных клеток – кардиомиоцитов (Рис. 14).
Перечислим особенности по сравнению со скелетными мышечными волокнами:
волокна имеют клеточное строение,
ядра клеток располагаются в средней части, а миофибриллы на периферии,
больше саркоплазмы и митохондрий и относительно меньше миофибрилл,
наличие вставочных пластинок. Это место стыка соседних кардиомиоцитов; к ним прикрепляются актиновые (тонкие) протофибриллы,
соседние мышечные волокна анастомазируются друг с другом, переходят друг в друга,
наличие волокон Пуркинье. Они образованы атипичными сердечными мышечными клетками; образуют проводящую систему сердца. Располагаются под эндокардом.
Особенности строения атипичных сердечных мышечных клеток:
большой диаметр,
мало миофибрилл и они располагаются на периферии,
в цитоплазме много гранул гликогена.
Сердечная мышечная ткань практически не восстанавливается на клеточном уровне в виду того, что в ее составе нет малодифференцированных клеток (источника регенерации).
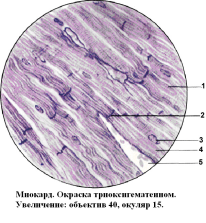
Рис. 14
2. Вставочный диск.
3. Ядра
5. Рыхлая соединительная ткань
ЛЕКЦИЯ 7
Нервная ткань
Нервная ткань (в составе нервной системы) осуществляет регуляцию деятельности тканей и органов и их связь с окружающей средой.
Нервная ткань состоит из нейронов, выполняющих основную функцию и клеток нейроглии, осуществляющих побочные (обслуживающую) функции по отношению к нейронам.
Основная функция нервной ткани слагается из следующих этапов:
1. Рецепция – это превращение энергии раздражителей в нервные импульсы. Рецепция осуществляется в рецепторах. Возникшие в них нервные импульсы далее передаются по нервным волокнам к нейронам.
2. Анализ и синтез поступающей информации. Эту, самую сложную часть нервной функции, выполняют нейроны в составе центральной нервной системы (головной мозг). В нейронах, поступающие из рецепторов нервные импульсы, анализируются, т.е. суммируются, ингибируются, или усиливаются. В результате этой сложной работы возникают качественно новые нервные импульсы, которые передаются рабочим органам и в результате мы имеем: или какое-то ощущение, или двигательную реакцию, или секрецию.
Нервная ткань развивается из утолщения эктодермы – нервной пластинки, которая затем замыкается в нервную трубку, имеющую следующие слои:
эпендимный, клетки которого дифференцируются в эпендимоциты,
плащевой – его клетки дифференцируются в нейробласты, а далее нейроны и спонгиобласты, из которых образуются клетки нейроглии,
краевая вуаль – из нее образуются нервные волокна
Нейроны или нейроциты.
Имеют следующие части:
тело (перикарион),
отростки,
концевые образования (окончания).
Отечественными нейрогистологами сформулирована нейронная теория строения нервной ткани, объясняющая связь нейронов, друг с другом. Она гласит, что нейроны функционально связаны, но анатомически изолированы, т.е. нейроны только контактируют друг с другом в области синапсов, но никогда непрерывно не переходят друг в друга.
Тело нейрона или перикарион – это ядросодержащая часть клетки. Ядро относительно крупное, светлое; характерно наличие крупного ядрышка. (Рис. 15)
В цитоплазме тела нейрона располагаются все общие органеллы. Характерно наличие и специальных органелл:
тельца Ниссля (базофильное вещество),
нейрофибриллы.
Тельца Ниссля выявляются при окрашивании основными красителями в виде гранул (глыбок). Под электронным микроскопом тельца Ниссля представляют из себя отдельные скопления компонентов (мембран и рибосом на их поверхности) гранулярной эндоплазматической сети, т.е. функционально – это места синтеза белковых соединений, которые далее транспортируются к отросткам и концевым образованиям.
Нейрофибриллы выявляются при окрашивании солями серебра в цитоплазме тела и отростков нейронов в виде нитевидных структур коричневого цвета. Под электронным микроскопом нейрофибриллы представляют из себя совокупность нейротрубочек и нейрофиламентов (аналоги немембранных органелл – микротрубочек и микрофиламентов). Функция – транспортная, т.е. вдоль нейрофибрилл транспортируются белковые и небелковые соединения, которые синтезируются в цитоплазме тела нейронов, в концевые образования.
Отростки нейронов.
Различают два вида отростков:
аксон
дендриты.
Определения. Аксон – это отросток нейрона, по которому нервные импульсы идут от тела нейрона. Нейроны имеют только один аксон. Дендриты – это отростки нейронов, по которым нервные импульсы идут к телу нейронов. Дендрит в составе нейронов может быть или один (если нейроны имеют только два отростка), или несколько. Цитоплазматическая мембрана отростков нейрона (оболочка) носит название – аксолема. А их цитоплазма – аксоплазма. В аксоплазме аксона имеются: нейрофибриллы, митохондрии и единичные рибосомы. В аксоплазме дендритов имеются: нейрофибриллы, митохондрии и компоненты гранулярной эндоплазматической сети (тельца Ниссля).
Классификация нейронов по количеству отростков. Различают нейроны:
униполярные (с одним отростком),
биполярные (с двумя отростками),
мультиполярные (со многими отростками) и
псевдоуниполярные.
Униполярных нейронов у человека нет; они имеются у низкоорганизованных животных (моллюсков, червей и т.д.).
Биполярные нейроныу человека имеются в составе: сетчатки глаза, вестибулярных и кохлеарных ганглиев.
Мультиполярные нейроны – это основной вид нейронов центральной нервной системы, т.е. в составе головного и спинного мозга.
В псевдоуниполярных нейронах спинномозговых узлов от тела отходит один отросток, который затем делиться на две ветви:
периферическая ветвь – это дендрит, оканчивается чувствительными нервными окончаниями в составе органов,
центральная – это аксон, вступают в контакт с нейронами центральной нервной системы.
По функциональной (физиологической) классификации различают следующие виды нейронов:
чувствительные,
двигательные,
ассоциативные.

Рис. 15
1. Тело нейрона
2. Плазмолемма
3. Аксоны и дендриты
Чувствительные нейроны связаны с чувствительными нервными окончаниями в составе всех органов. Они образуют начальное (чувствительное) звено рефлекторной дуги любого рефлекса (соматического и вегетативного). Пример. Псевдоуниполярные нейроны спинномозговых узлов функционально чувствительные.
Двигательные нейроны связаны с двигательными нервными окончаниями в составе разных органов. Они образуют конечное (двигательное) звено рефлекторной дуги. Пример. Двигательные нейроны передних рогов спинного мозга.
Ассоциативные нейроны располагаются между чувствительными и двигательными; образуют промежуточное (ассоциативное) звено рефлекторной дуги. Пример. Большая часть нейронов центральной нервной системы функционально ассоциативные.
Нейроглия. Они создают стабильную внутреннюю среду для нервной ткани, выполняя: опорную, разграничительную, трофическую, секторную, защитную и другие функции.
Классификация нейроглии.
Включает в себя: макроглию и микроглию. В понятие – макроглия входят:
эпиндимная глия,
астроцитная глия и
олигодендро глия
Эпендимная глия. Клетки имеют коническую форму. На одной (широкой) поверхности располагаются реснички; с противоположной поверхности отходит один отросток, который затем ветвится. Эти клетки выстилают стенку спинномозгового канала и желудочков мозга в виде одного слоя клеток. Функции:
способствуют циркуляции спинномозговой жидкости колебаниями своих ресничек,
в цитоплазме этих клеток образуются некоторые химические компоненты спинномозговой жидкости.
Астроцитная глия
Различают астроциты:
Протоплазматические астроциты имеют короткие, толстые, сильно ветвящиеся отростки. Располагаются в сером веществе мозга.
Волокнистые астроциты имеют длинные, тонкие, слабо ветвящиеся отростки. Располагаются в белом веществе мозга.
Функции астроцитов:
опорная. Располагаясь между нейронами удерживают их в определенном положении,
разграничительная (изолируют нейроны друг от друга),
трофическая. Нейроны не имеют связи с капиллярами. Они получают питательные вещества только через цитоплазму астроцитов, которые одними своими отростками связаны с капиллярами, а другими – с телами нейронов.
Олигодендроглия. Это мелкие клетки с короткими ветвящимися отростками. Они плотно прилежат к телам и отросткам нейронов, образуя оболочки. Функции те же, что у астроцитов.
Микроглия.
Источником ее развития является
мезенхима. Это клетки малых размеров с
многочисленными короткими отростками.
Функционально – это макрофаги нервной ткани (Рис. 16)..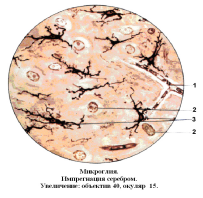
Рис. 16
Нервное волокно
Ядра нейронов
Микроглиоциты
механизм мышечного сокращения
Книга «Болезни сердечно-сосудистой системы (Р.Б. Минкин)».
Механизм мышечного сокращения
Мышца преобразует химическую энергию непосредственно в механическую энергию (работу) и теплоту. Сокращение мышцы при постоянной нагрузке называется изотоническим, при постоянной длине — изометрическим.
Источником энергии для сокращения служит АТФ. Во время сокращения АТФ расщепляется с помощью гидролиза до аденозиндифосфата (АДФ) и неорганического фосфата (Pi): АТФ -АДФ + Pi.
Восстановление АТФ происходит с помощью расщепления углеводов и распада креатинфосфата (КФ): КФ + АДФ — АТФ + К (К-креатин). АТФ расщепляется и энергетически утилизируется в мышце с помощью фермента миозина — АТФазы.
Этот процесс активизируется актином в присутствии ионов магния. Головки миозина, которые взаимодействуют с актином, содержат активные каталитические центры для расщепления АТФ.
Поэтому АТФ расщепляется только в случае прикрепления головки миозина к активирующему белку — актину, при этом образуются актомиозиновые мостики.
Сокращению мышцы предшествует ее возбуждение. Возбуждение, деполяризация, происходит под влиянием потенциала действия, поступающего через нервно-мышечные синапсы.
Передача сигнала о сокращении от возбужденной мембраны кардиомиоцита к миофибриллам в глубину клетки называется электромеханическим сопряжением.
В электромеханическом сопряжении ключевая роль принадлежит ионам Са2 +. Проникновение возбуждения вглубь мышечного волокна с его поверхности происходит с помощью поперечных Т-трубочек. Мембрана этих трубочек обладает высокой возбудимостью и способностью проводить возбуждение.
Они играют важную роль в процессе передачи сигнала от клеточной мембраны к хранилищам кальция в клетке. При этом кальций высвобождается из хранилищ в продольной тубулярной системе.
В расслабленном состоянии концентрация ионов Са2 + в клетке примерно в 10 000 раз меньше, чем во внеклеточном пространстве. Хранение и высвобождение ионов Са2 + происходит из системы поперечно-продольных трубочек. В поперечные трубочки клетки Са2 + поступает из внеклеточного пространства, с которым эти трубочки связаны.
Продольные трубочки не связаны с внеклеточной средой, и Са2 + хранится в их концевых разветвлениях — цистернах, откуда он и поступает внутрь клетки при ее возбуждении. Возбуждение, проникшее внутрь клетки, приводит к высвобождению ионов Са2 + из цистерн во внутреннюю среду клетки около миофибрил, что и приводит к их сокращению.
При расслаблении ионы Са2 + удаляются посредством кальциевого насоса в систему каналу саркоплазматического ретикулума. Снижение концентрации Са2 + подавляет активность АТФазы актомиозина и происходит разъединение нитей актина и миозина.
При расслаблении миофибрилл во время диастолы в отсутствие ионов Са длинные молекулы тропомиозина располагаются так, что они прикрывают собой активные центры актиновых нитей и тем самым не дают возможности образоваться связи между актином и миозином.
Актомиозиновые мостики не образуются. Ионы Са2 + , поступающие при возбуждении в саркоплазму клетки, образуют с тропонином Са2 + — тропониновые комплексы.
При этом изменения, возникшие в молекуле тропонина, приводят к смещению тропомиозина и открытию активных центров в актиновых нитях (рис.8). К активным центрам присоединяются головки молекул миозина. Возникшее соединение — актомиозиновый мостик — с помощью «гребущих» движений приводит к скольжению нитей актина и миозина друг относительно друга и укорочению саркомера на 25 — 50 %.
Сами же нити актина и миозина при таком скольжении не укорачиваются. Такой механизм сокращения называется моделью скользящих нитей и был предложен Хаксли в 50-х годах. Мышечная сила развивается за счет энергии АТФ.
Сила и скорость сокращений регулируется количеством открытых актиновых центров, числом образованных актомиозиновых мостиков, возможностями восстановления, ресинтеза, АТФ в митохондриях и рядом других факторов.
В каждом цикле прикрепления—отсоединения актомиозинового мостика АТФ расщепляется только один раз. Чем больше мостиков находится в активном состоянии, тем выше скорость расщепления АТФ и сила, развиваемая мышцей.
Мышечное сокращение происходит тем быстрее, чем скорее перемещается актомиозиновый мостик, т. е. чем больше «гребущих» движений происходит в единицу времени. Когда движение мостика завершается, с ним связывается новая молекула АТФ, и начинается новый цикл. Координированное сокращение всех миофибрилл приводит к сокращению сердечной мышцы — систоле сердца. Отсоединение актомиозиновых мостиков приводит к расслаблению мышцы — диастоле сердца.
Сердце как насос. Насосная, механическая, или сократительная, функция сердца обеспечивает движение крови по сосудистой системе организма. Вильям Гарвей в 1628 г. впервые показал, что сердце нагнетает кровь в сосуды. У человека в состоянии покоя во время каждой систолы желудочки сердца выбрасывают 70 — 80 мл крови, так называемый ударный объем (УО) — левый желудочек — в аорту, правый — в легочную артерию.
Сокращаясь при этом 65 — 75 уд/мин, оно выбросит приблизительно около 5 л крови, что называется минутным объемом (МО). Каждый сердечный цикл при такой частоте ритма продолжается примерно 0,8 с; из них 0,3 с приходится на период сокращения, систолу, и 0,5 с — на период расслабления, диастолу.
Работа, совершаемая при этом сердцем, очень велика. Она равна произведению массы крови, выбрасываемой с каждой систолой на сопротивление в сосудах (в аорте — для левого желудочка и в легочной артерии — для правого).
Такая работа, производимая сердцем в течение суток, составляет приблизительно 216 кДж и эквивалентна силе, достаточной для подъема груза массой 2,2 кг из самой глубокой морской впадины на самую высокую гору. Сердце в сутки в среднем 9 ч работает и 15 ч отдыхает. При нагрузке
механизм мышечного сокращения
за счет увеличения частоты и силы сокращения сердце может увеличить выброс крови с 5 до 25 л/мин. Правая и левая половины сердца (соответствующие предсердие и желудочек) составляют как бы два насоса. Предсердия и желудочки соединены фиброзными кольцами атриовентрикулярных клапанов, и пучок Гиса является единственной мышечной связью между ними.
При повышении давления в предсердиях выше давления в желудочках атриовентрикулярные клапаны открываются, и кровь поступает из предсердий в желудочки. Во время систолы желудочков атриовентрикулярные клапаны закрываются, и это препятствует обратному поступлению крови, регургитации (англ. regurgitate — хлынуть обратно), из желудочков в предсердия.
Выворачиванию атриовентрикулярных клапанов в сторону предсердий препятствует натяжение прикрепленных к ним сухожильных хорд папиллярных мышц. Полулунные клапаны аорты и легочной артерии открываются во время изгнания крови из соответствующего желудочка и закрываются, когда давление крови в сосуде становится выше, чем давление в желудочке.
После систолы желудочка в нем может оставаться небольшое Количество крови, которое называется конечно-систолическим объемом (КСО). В связи с тем, что давление, развиваемое левым желудочком из-за высокого давления в аорте, примерно в 5 раз больше , чем в правом, работа левого желудочка в 5 раз превышает работу правого желудочка.
Во время сокращения сердце поворачивается в грудной клетке таким образом, что его верхушка приближается к грудной стенке в межреберном промежутке, образуя «верхушечный толчок».
В начале нынешнего столетия Уиггерс впервые произвел синхронную запись изменений давления крови в предсердиях и желудочках сердца, а также в крупных сосудах, отходящих от сердца, и электрических и звуковых процессов, возникающих при работе сердца.
Это позволило ему в 1921 г. произвести разделение сердечного цикла на отдельные базы. Такое разделение с некоторыми уточнениями является общепринятыми на сегодняшний день, оно позволяет оценить сократительные свойства миокарда (рис.9).
Начальную часть систолы желудочка называют электромеханическим латентным интервалом. Ему соответствует период времени между началом зубца Q на ЭКГ и низкочастотными колебаниями I тона на ФКГ. В это время происходит распространение возбуждения по миокарду желудочков. Отдельные волокна начинают сокращаться, однако их число недостаточно для систолы всего желудочка.
Следующий этап систолы желудочка называется фазой асинхронного сокращения. Эта фаза продолжается от начала подъема давления в желудочке до начала высокочастотных колебаний I тона на ФКГ. В этот период происходит последовательное сокращение различных участков миокарда желудочка.
Но поскольку сокращение протекает неравномерно, асинхронно, то повышения давления в желудочке практически не происходит. Давление повышается в следующую фазу систолы — изометрического, или изоволюмического, сокращения (греч. isos — равный, англ. volume — объем). В этот
механизм мышечного сокращения
период давление крови в желудочке повышается сначала медленно, затем — очень быстро. Во время этой фазы атриовентрикулярные клапаны уже закрыты, а полулунные — еще не открылись.
Поскольку кровь, как всякая жидкость, несжимаема, то сокращение желудочков происходит при неизменном объеме.
Энергия сокращения преобразуется в энергию давления. Давление в желудочках повышается практически от нуля до уровня давления в аорте к концу диастолы (примерно 80 мм рт. ст.) — в левом желудочке и до уровня давления в легочной артерии (примерно 10—15 мм рт. ст.) — в правом желудочке.
Когда давление в желудочках достигает уровня давления в крупных сосудах, исчезает градиент (разница) давления и происходит открытие полулунных клапанов аорты и легочной артерии — протосфигмический интервал. Открытие клапанов происходит за 0,01 — 0,02 с.
Эти этапы систолы соответствуют периоду напряжения желудочков, которые подготавливают их к изгнанию крови. Изгнание крови происходит в 2 фазы: фазу максимального и фазу замедленного изгнания, или редуцированного изгнания. В первую фазу желудочек выбрасывает примерно Уз систолического объема крови, во вторую — Уз.
Во время фазы максимального изгнания давление в желудочках и крупных сосудах продолжает увеличиваться, достигая максимальных значений для левого желудочка примерно 120 мм рт. ст., для правого — 25 мм рт. ст. В это время объем желудочков резко уменьшается.
Отток крови через ветви аорты и легочной артерии в течение фазы замедленного изгнания превышает ее поступление в сосуды, поэтому давление в желудочках и крупных сосудах снижается.
Суммарная длительность периодов напряжения и изгнания составляет продолжительность так называемой электромеханической, или общей систолы; время изометрического сокращения и периода изгнания соответствует механической систоле сердца. Во время механической систолы в желудочке нарастает и удерживается высокое давление крови. После этого начинается диастола.
Диастола начинается протодиастолическим интервалом, в течение которого закрываются полулунные клапаны аорты и легочной артерии. Теперь, когда полулунные клапаны уже закрыты, а предсердно-желудочковые еще не открылись, давление в желудочках быстро снижается до уровня давления в предсердиях.
Это время соответствует фазе изометрического, или изоволюмического, расслабления. Пока желудочки сокращались, предсердия находились в состоянии диастолы и наполнялись кровью, поэтому давление в них постепенно повышалось.
Суммарная длительность протодиастолического интервала и фазы изометрического расслабления соответствует длительности периода расслабления желудочков.
При снижении давления в желудочках до уровня давления в предсердиях открываются предсердножелудочковые клапаны, и желудочки начинают наполняться кровью. Сначала в силу максимальной разницы, градиента, давления — относительно высокое в предсердиях и низкое в желудочках, наступает фаза быстрого наполнения желудочков кровью.
Затем давление в полостях сердца выравнивается и наступает фаза медленного наполнения, или диастазис, которая заканчивается систолой предсердий.
В период диастолы объем желудочков увеличивается. При замедлении предсердно-желудочковой проводимости между окончанием систолы предсердий и началом систолы желудочков иногда выделяют интерсистолический интервал.
Фазы сердечного цикла эквивалентны для обеих половин сердца. Ниже представлены данные о длительности фаз у здоровых лиц (В. Л. Карпману).
механизм мышечного сокращения
Необходимо учитывать зависимость отдельных фаз от частоты ритма сердечных сокращений. Для этого сравнивают реально найденное значение с должной величиной, рассчитанной для данного ритма:
Е = 0,109 хС + 0,159 и Sm = 0,114 хС + 0,185,
где Е — длительность периода изгнания; С — длительность сердечного цикла; Sm — длительность механической систолы.
Изменение длительности фаз сердечного цикла происходит при нарушении сократительных свойств миокарда, но может зависеть и от внесердечных причин, нарушающих работу сердца (например, от повышения артериального давления и др.).
Франк и независимо от него Старлинг показали, что при увеличении диастолического наполнения сердца увеличивается ударный выброс крови (УО). Увеличение УО происходит за счет увеличения силы сердечного сокращения. Сердце выполняет повышенную работу за счет увеличения начальной длины волокон миокарда при увеличении диастолического наполнения желудочков.
Таким образом, по закону Франка—Старлинга механическая энергия, освобождающаяся при переходе мышцы из состояния покоя в состояние сокращения, зависит от исходной длины мышечного волокна. Сила сокращения тем больше, чем сильнее растянуты ее волокна.
Такой параллелизм между силой сердечных сокращений и степенью растяжения мышечных волокон наблюдается лишь до определенных пределов, пока сохраняется нормальным тонус миокарда.
Полагают, что в основе механизма закона Франка—Старлинга лежит увеличение связывания ионов Са + с тропонином в процессе сокращения миофибрилл.
На сократимости миокарда сказываются, помимо закона Франка-Старлинга, нервные воздействия. Раздражение симпатических нервных окончаний, как и повышение концентрации в крови катехоламинов, увеличивает силу сердечных сокращений без увеличения начальной длины волокон миокарда. Волокна блуждающего нерва не оказывают заметного влияния на сократительную способность миокарда желудочков.
МО при умеренной мышечной работе увеличивается с 5 до 12 — 15 л, при усиленной — до 20 — 25 л. Увеличение МО происходит за счет УО и частоты сердечных сокращений. Это сопровождается уменьшением КСО и увеличением конечно-диастолического (КДО) объема крови в желудочках сердца.
Укорачивается систолический и особенно диастолический интервалы желудочков, резко возрастает потребление миокардом кислорода.
У спортсменов, в отличие от нетренированных лиц, УО сердца больше как в покое, так и особенно при нагрузке. Это обусловлено физиологической гипертрофией миокарда и увеличением объема сердца. Поэтому у спортсменов нагрузка сопровождается главным образом увеличением УО без существенного нарастания частоты сердечных сокращений, в то время как у нетренированных лиц такая же нагрузка вызывает, наоборот, резкое увеличение частоты без существенного увеличения УО.
Такая реакция миокарда на нагрузку является значительно менее энергетически целесообразной. Энергетика миокарда была изучена в 50-х годах Бингом с помощью катетеризации коронарного синуса сердца. Процессы, связанные с выработкой энергии, универсальны для всего живого, но освобождение энергии в разных органах и у разных видов происходит по-разному.
Исходные пищевые вещества — углеводы, белки и жиры — расщепляются в организме до очень простого соединения — уксусной кислоты, которая в дальнейшем превращается в так называемую «активную уксусную кислоту». Активная уксусная кислота включается в процесс, связанный с выработкой энергии (цикл Кребса). Этот цикл составляет биохимическую основу клеточного дыхания.
В результате процессов, идущих с поглощением кислорода (аэробного окисления), в этом цикле образуются молекулы высокоэнергетического фосфорного соединения АТФ. АТФ является источником энергии для сокращения миокарда. Скорость обмена АТФ в миокарде, как и его синтез, очень высокие.
Работающее сердце постоянно нуждается в кислороде и максимально извлекает его из крови коронарных артерий. Единственный способ, с помощью которого сердце покрывает повышенную потребность в кислороде при нагрузке, является увеличение коронарного кровотока. Потребление кислорода пропорционально напряжению, развиваемому миокардом. Обмен веществ в миокарде почти целиком идет с поглощением кислорода, т. е. аэробен.
Потребление кислорода миокардом в состоянии покоя составляет около 25 %. При сужении или закупорке коронарной артерии кровоток через нее не может возрастать, возникает кислородная недостаточность, ишемия миокарда. Это сопровождается симптомами коронарной недостаточности (стенокардия, инфаркт миокарда).
Сердце в процессе обмена веществ использует большие количества углеводов, жирных кислот, кетоновых тел, аминокислот и других субстратов. Большую часть необходимой энергии миокард получает за счет обмена жирных кислот и углеводов.
Свободные жирные кислоты переносятся в ионизированной форме через клеточную мембрану с помощью диффузии. Внутри кардиомиоцита они связываются со специальным белком. При увеличении работы сердца скорость поглощения клеткой свободных жирных кислот возрастает, при этом ускоряется расщепление, гидролиз, АТФ. Глюкоза поступает в кардиомиоцит через его наружную мембрану с помощью специального переносчика.
Скорость поглощения глюкозы клеткой увеличивается под действием инсулина и при увеличении совершаемой сердцем работы. В клетке молекулы глюкозы объединяются в полисахарид—гликоген. Гликоген постоянно участвует во внутриклеточном обмене веществ, он служит потенциальным источником энергии, так как способен распадаться на отдельные молекулы глюкозы (гликогенолиз).
Коэффициент полезного действия сердца, который определяется по отношению совершенной работы к затраченной энергии, составляет всего 15 — 25 %. Остальная часть энергии рассеивается в основном в виде тепла (до 50 %).
Особенности сократимости сердечной мышцы
Зависимость “сила стимула- сила сокращения”
В отличие от скелетной мышцы сила сокращения сердечной мышцы не зависит от силы раздражителя закон “всё или ничего”. В опыте изолированное сердце лягушки на допороговое раздражение вообще не отвечает, но как только сила раздражения достигает порогового уровня, возникает его максимальное сокращение (рис.5).
Дальнейшее увеличение силы раздражающего тока не изменяет величины сокращения. Подчинение сердечной мышцы закону “всё или ничего” объясняется особенностями строения миокарда, клетки которого образуют функциональный синцитий: все мышечные клетки соединены друг с другом вставочными дисками с очень низким электрическим сопротивлением и в функциональном плане представляют собой единое образование. Поэтому пороговый раздражитель приводит к возбуждению сразу всех кардиомиоцитов и развитию максимального сокращения.

Рис. 5. Независимость силы сокращений миокарда (а) от силы раздражителя (б) – закон «все или ничего». Пороговый стимул отмечен звездочкой.

Рис.6. Зависимость силы сокращений миокарда (а) от частоты стимуляции (б) – «лестница Боудича», полученная на сердце лягушки, предварительно остановленном с помощью первой лигатуры Станниуса.
Зависимость “частота-сила”.
Закон “всё или ничего” для миокарда не абсолютен. Если в эксперименте раздражать мышцу желудочков импульсами возрастающей частоты, не меняя их силы, то величина сокращения миокарда будет возрастать на каждый следующий стимул (лестница Боудича или хроноинотропный эффект). Объясняется такой эффект тем, что при переходе к более высокой частоте стимуляции промежутки времени между сокращениями укорачиваются, вследствие чего не происходит полного удаления ионов кальция, поступивших в клетку при очередном сокращении. В результате с каждым последующим сокращением концентрация внутриклеточного кальция возрастает и соответственно возрастает и сила сокращения (рис 6).
Возбудимость сердечной мышцы во время сокращения.
Для изучения возбудимости надо наносить раздражение электрическим током пороговой или сверхпороговой силы на сердце лягушки в разные фазы его цикла. При этом сердце не ответит на раздражение, если оно будет нанесено в период систолы, когда миокард находится в состоянии абсолютной невозбудимости, т.е. рефрактерности (рис.11). Обратите внимание, что рефрактерный период занимает всю систолу и начало диастолы (рис.7). С началом расслабления возбудимость миокарда начинает восстанавливаться, и наступает фаза относительной рефрактерности.

Рис. 7. Графики сокращения, потенциала действия и возбудимости мио карда желудочков.
Экстрасистола желудочков. Нанесение сверхпорогового раздражения в фазу относительной рефрактерности способно вызвать внеочередное сокращение желудочков экстрасистолу. При этом пауза, следующая за желудочковой экстрасистолой, длится дольше, чем обычная, так называемая компенсаторная пауза. Большая длительность этой паузы объясняется тем, что очередной импульс из синусного узла застаёт желудочки в период рефрактерности уже полученной экстрасистолы, и нормальное их сокращение возможно только с приходом очередного импульса (рис.8).
У человека дополнительные, внеочередные импульсы, вызывающие экстрасистолу, могут возникать в норме в элементах проводящей системы или в самом миокарде желудочков при активации симпатического отдела вегетативной нервной системы (например при эмоциональном возбуждении), а также при патологических процессах в миокарде.
Итак, абсолютная невозбудимость миокарда, продолжающаяся всю систолу, делает сердце нечувствительным в этот период к дополнительным раздражениям, исключает возможность длительного непрерывного (тетанического) сокращения, и тем самым помогает сердцу работать в режиме одиночного сокращения. Длительная рефрактерность гарантирует продолжение диастолы даже при возникновении внеочередных раздражений, и создаёт условия для наполнения желудочков кровью, т.е. для поддержания минутного объёма сердца.
Рефрактерность кардиомиоцитов обеспечивает также нормальную последовательность распространения возбуждения в сердце, препятствует возникновению кругового движения возбуждения по миокарду.

рис.8. График желудочковой экстрасистолы
Стрелками отмечен момент нанесения внеочередного раздражения, треугольничками момент поступления очередного импульса из синоатриального узла.
Синусовая экстрасистола. При эмоциональном возбуждении или под влиянием воспалительных изменений внеочередной импульс возбуждения может возникнуть в самом синусном узле, следствием которого будет полный внеочередной цикл сердца, за которым в отличие от желудочковой экстрасистолы не следует компенсаторная пауза. Понятно, что пауза перед внеочередным сокращением будет укорочена (рис. 9).

Рис.9. Синусовая экстрасистола (обозначена стрелочкой).
2. Проводимость сердечной мышцы: понятие, механизм, особенности.
Под проводимостью сердечной мышцы подразумевается процесс распространения электрических потенциалов, самопроизвольно возникающих в определенных сердечных клетках.
Сердце состоит из двух основных групп сердечных клеток: клетки рабочего миокарда, основная роль которых заключается в ритмических сокращениях, обеспечивающих насосную функцию сердца, и клетки проводящей системы. Проводящая система состоит из: 1)синусового узла, находящегося в правом предсердии; 2)атриовентрикулярного узла, находящегося на границе предсердий и желудочков; 3)непосредственно проводящей системы, включающей пучок Гисса, расположенный на границе желудочков и переходящий в левую и правую ножки и волокна Пуркинье, пронизывающие клетки рабочего миокарда желудочков.
Одна из главных особенностей сердечной мышцы — это наличие особых контактов между ее клетками. Эти контакты образованы участками мембран прилегающих соседних клеток и, благодаря их особым свойствам (в частности, низкому сопротивлению, в то время как мембрана кардиомиоцита вне зоны контакта обладает высоким сопротивлением), позволяют электрическому току распространяться от клетки к клетке. Поэтому сложноустроенная сердечная мышца при сокращении ведет себя почти как одна гигантская клетка.
Роль клеток проводящей системы заключается в генерации возбуждения, то есть в ритмической генерации импульсов электрического тока специфической формы и величины. Эти импульсы исходно возникают в синусовом узле, распространяются по проводящей системе в атриовентрикулярный узел и оттуда идут по пучку Гисса и волокнам Пуркинье, достигая клеток рабочего миокарда и вызывая их ритмические сокращения.
3. Задача. У человека установлена задержка оттока желчи из желчного пузыря. Влияет ли это на переваривание жиров?
Преобладающими в желчи веществами являются соли желчных кислот, составляющие около половины общего количества растворенного вещества в желчи. Желчные соли выполняют в кишечном тракте две важные функции. Во-первых, они обладают детергентным свойством в отношении жировой части пищи. Это проявляется в снижении поверхностного натяжения частиц пищи, что позволяет дробить частицы жира на мелкие части при перемешивании в кишечнике. Этот процесс носит название эмульгирования, или детергентной функции желчных солей.
Во-вторых, наиболее важной после детергентной функции является способность желчных солей помогать всасыванию: (1) жирных кислот; (2) моноглицеридов; (3) холестерола; (4) других липидов из кишечного тракта. Они осуществляют это, образуя очень малые физические комплексные соединения с липидами. Эти соединения называют мицеллами, они наполовину растворимы в химусе из-за электрических зарядов желчных солей. Кишечные липиды «транспортируются» в такой форме к слизистой кишечника, где затем всасываются в кровь. При отсутствии желчных солей в кишечном тракте с калом теряется до 40% принятых внутрь жиров, и у человека часто развивается метаболический дефицит, обусловленный потерей этих питательных веществ.
Билет №11
Физиология мышечного сокращения: работа мышц человека

Для студентов спортивных вузов, атлетов и всех любознательных объясняем простым языком источники энергии и физиологию (механизм) мышечного сокращения, а также строение мышц, с приведением простейших формул и фото, понимание которых, вам поможет в дальнейшим более эффективно тренироваться в тренажерном зале.
В теле человека находиться более 600 мышц, и каждая из них отвечает за тот или иной вид деятельности, например, мышцы спины обеспечивают ровную осанку, удерживают тело в вертикальном положении, а благодаря глазодвигательным мышцам, мы можем направлять свой взгляд в рассматриваемом направлении.
С помощью мышц мы познаем окружающий мир, окружающую действительность, если бы не они, мы бы так и не научились ходить, писать, говорить, выражать свои эмоции. Однако, для того чтобы мышцы нормально функционировали, их необходимо снабжать энергией, полезными, питательными веществами, если в вкратце, то процесс пищеварения выглядит следующим образом:
После механического перемалывания во рту пища через глотку попадает в пищевод, далее в желудок, там под действием желудочного сока (в основном соляная кислота и фермент пепсин) происходит расщепление белков и частично жиров, и попадает в привратник, далее пищевая масса переходит в двенадцатиперстную кишку (начало тонкой кишки), где под действием поджелудочного сока и желчи происходит дальнейшее разложение белков, углеводов и жиров, далее химус переходит в тонкий кишечник, в котором происходит окончательное разложение аминокислот, жирных кислот и моносахаридов и благодаря наличию в нем ворсинок всасываются в стенки тонкого кишечника, все что не всосалась, переходит в толстый кишечник, в котором начинают иди процессы размельчения клетчатки и химуса, а также всасывание остаточной воды и формироваться кал.
Всасывая питательные вещества через желудок и кишечник, полученные вещества попадают в кровь (в воротную вену), и далее направляться в печень, после чего попадают в общий кровоток омывая все ткани человека.
Мышечная система человека
Мышечная система человека позволяет координировать движения тела, держать его в равновесии, осуществлять дыхание, а также транспорт пищи и крови внутри организма, помимо всего она защищает внутренности от повреждений, а также выполняет роль преобразователя энергии химической в механическую и тепловую.
В теле человека всего три типа мышц:
- скелетные
- гладкие
- мышца сердца
 Мышечная система человека (A — мышца сердца, B — скелетные мышцы, C — гладкие мышцы)
Мышечная система человека (A — мышца сердца, B — скелетные мышцы, C — гладкие мышцы)Скелетная мускулатура
Скелетная мускулатура человека, она же поперечнополосатая, крепится к костям, состоит из волокон, а они в свою очередь состоят из мышечных клеток. В каждой мышечной клетке имеется два ядра, которые отвечают за деление и восстановление. За сокращение мышцы отвечают, так называемые миофибриллы (нити), которые содержаться в мышечных клетках. Количество миофибрилл в мышечной клетке может достигать до несколько тысяч. Таким образом, мышечные клетки формируют ткань, а она в свою очередь образовывает мышцу.
Наши скелетные мышцы содержат волокна, нервные окончание и кровеносные сосуды. Сокращение мышцы происходит с помощью нервных импульсов, которые поступают от спинного мозга до мышечной ткани, то есть передача нервного импульса осуществляется по пути — головной мозг → спинной мозг → нужные нам мышцы. Теперь понятно, почему повреждение спинного мозга так опасно.
Человек регулирует интенсивность сокращения мышц с помощью силы подаваемого импульса по нервным окончаниям.
 Скелетная мускулатура человека
Скелетная мускулатура человекаГладкие мышцы
Гладкая мускулатура выполняет не произвольные сокращения, состоит из веретеновидных клеток, являясь одной из самых важных составляющих мышечных полых органов, а также составной частью кровеносных и лимфатических сосудов, помогает транспортировать содержимое полых органов (транспорт пищи кишечнику), сужения зрачка, корректировка артериального давления, и другие процессы, которые происходят непроизвольно.
Все сокращения гладким мышц не вызывают утомления, регулируются вегетативной системой (автономная нервная система, которая отвечает за работу внутренних органов).
Натренировать гладкие мышцы можно, например, увеличивая выносливость, вы улучшаете работу сердечно-сосудистую системы.
 Гладкие мышцы
Гладкие мышцыСердечная мышца
Сердце непрерывно сокращается в течении всей жизни, обеспечивая движение, перекачку крови, питательных веществ, других жизненно-важных веществ по сосудам к тканям организма. Выполняя роль насоса, сердце работает в режиме непрерывных, ритмичных, одиночных сокращений.
Строение волокна миокарда, напоминает структуру скелетных мышц, которые также содержат миофибриллы, состоящие из актина и миозина, включая тропонин-тропомиозиновый белковый комплекс.
Картинку сердце, где показанна устройство сердца где можно увидеть миокард
Механизм мышечного сокращения сердца, происходить все по тем же причинам, что и в поперечнополосатых мышцах, благодаря ионами Ca2+ (кальция), которые освобождаются из саркоплазматического ретикулума (мембранная органелла мышечных клеток), только в этом случае он менее упорядочен (по сравнению со скелетной мускулатурой).
 Сердечная мышца и ее устройство
Сердечная мышца и ее устройствоСтроение мышц (мышечных волокон) человека
Мышцы человека состоят из мышечных волокон, которые в свою очередь состоят из мышечных клеток. Взятое в отдельности мышечное волокно представляет собой многоядерную мышечную клетку, диаметр которой варьируется от 10 до 100 мкм, которая имеет оболочку сарколемму (клеточная мембрана), заполненной саркоплазмой (содержимое клетки, основа которой — матрикс). Миофибриллы располагаться в саркоплазме, то есть саркоплазма заполняет пространство между миофибриллами и окружает ядра клеток. Миофибрилла представляет собой нитевидной формы образование, состоящее из саркомеров (сократительный аппарат мышцы).
 Строение скелетной мышцы
Строение скелетной мышцыВ зависимости от количества миофибрилл, различают белые и красные мышечные волокна.
Белые мышечные волокна отличаются от красных, большим количеством миофибрилл, и меньшим саркоплазмы, такое соотношение обеспечивает быстроту сокращение белых волокон. Благодаря наличию миоглобина (кислородосвязывающий белок) в мышцах, который придает цвет, мышечные волокна называют красными.
Саркоплазма в мышечных клетках содержит помимо миофибрилл, еще и митохондрии (энергетические станции клеток, в которых синтезируются АТФ), рибосомы, комплекс Гольджи, жировые включения, и другие постоянные компоненты клетки, без которых существование ее не возможно (органоиды).
Передача импульсов возбуждения внутри мышц происходит благодаря саркоплазматической сети. Базовая сократительная единица поперечнополосатых мышц (саркомеры) в своем составе содержат толстые миозиновые нити, образованные белком миозином, и тонкие актиновые нити, образованные белком актином.
Актин – сократительный белок, на который приходиться около 15% от всего мышечного белка, содержится в тонких филаментах скелетных мышц, обеспечивая осуществление двигательных функций клеток.
Миозин – основной белок, из которого состоят мышечные волокна, благодаря которому мышцы имеют эластичность и способны сокращаться. Масса миозина составляет порядка 55% от всех сократительных белков, которые содержаться в мышечных волокнах.
Миозин сконцентрирован в поперечнополосатых мышцах (скелетной мускулатуре), которые отвечают за рефлексы и целенаправленность движений. Благодаря способности миозина расщеплять АТФ химическая энергия макроэргических связей АТФ переходит в механическую энергию мышечного сокращения.
 Строение мышц человека
Строение мышц человекаАктомиозин – комплекс, состоящий из белков актина и миозина, создает мышечные волокна, которые распределяются в определенном порядке. Сокращение актомиозина возможно, благодаря энергии, которая освобождается в результате взаимодействия АТФ с водой (гидролиз), таким образом, актомиозин определяет способность мышц к сокращению (мышечное сокращение).
Механизм мышечного сокращения
Мышечное сокращение происходит благодаря, скольжению актиновых нитей вдоль миозиновых (скользящее сокращение), при этом сокращается общая длина актомиозинового комплекса, без изменения длины нитей актина и миозина. Первостепенную роль в сокращении мышц играют временно замыкающиеся поперечные мостики (головки миозиновых молекул), которые и обеспечивают продвижение актиновых нитей вдоль миозиновых. Соответственно, сила мышечного сокращения будет завесить от мостиков, — чем больше прикреплено мостиков к актиновым нитям, тем сильнее сокращение.
Энергия, необходимая для такова «скользящего сокращения», обеспечивается взаимодействием АТФ с актомиозином, в результате аденозинтрифосфорная кислота (АТФ), распадается на аденозиндифосфат (АДФ) и фосфорную кислоту (h4PO4).
Наряду с АТФ, важную роль в сокращении мышечного волокна играют еще ионы кальция, магния и вода.
Количество воды, которая содержит мускулатура, равняется примерно 72-80%, при этом мышечная ткань также содержит большое количество белков, и в малых количествах — гликоген, фосфолипиды, холестерин, креатинфосфат, креатин, витамины.
Чем больше мышечных волокон в скелетных мышцах, тем сильнее они будут.
 Механизм мышечного сокращения человека
Механизм мышечного сокращения человекаЭнергия мышечных сокращений
Как мы уже знаем, сокращение мышц, это движение (размыкание-смыкание) поперечных мостиков, и естественно, это процесс нуждается в энергообеспечении. Однако, запас энергии в мышцах очень мал, поэтому ее необходимо постоянно откуда-то «черпать», восстанавливать.
Основным и главным поставщиком энергии, благодаря которому происходит энергообеспечение организма, служит молекула АТФ, которая расщепляется, благодаря головкам миозиновых мостиков, таким образом, образуется энергия для сокращения мышц. Однако, мышечная клетка содержит в себе ограниченный запас молекул АТФ, их количество хватает всего на 8 повторений, первые 2 секунды:
- АТФ + h3O = АДФ + h4PO4 + энергия
Проблема в ограниченном обеспечении энергии для мышечного сокращения легко решается с помощью ресинтеза или пересоздания молекул АТФ.
 АТФ — как универсальный источник энергии человека
АТФ — как универсальный источник энергии человекаИсточники энергии мышц
В зависимости от времени нахождения мышц под нагрузкой, а также от степени интенсивности выполнения того или иного упражнения, можно выделить следующие виды систем, которые обеспечивают энергию для мышц:
- Фосфогенная (АТФ и КрФ)
- Гликолитическая (анаэробный и аэробный гликолиз)
- Окисление (расщепляются углеводы и липиды при активном поступлении кислорода)
В отдельных случаях, при длительном отсутствии потребления углеводов и жиров, могут в качестве источника энергии выступать белки (аминокислоты), благодаря процессу глюконеогенеза, при котором происходит превращение аминокислот в пируват, суммарная формула ниже:
2 Пируват + 4ATP + 2GTP + 2NADH + 2H+ + 4H2O → глюкоза + 4ADP + 2GDP + 6Pi + 2NAD+
Каждая энергетическая система характеризуется мощностью и емкостью.
Мощность – показатель, который ограничивает интенсивность выполняемой физической работы, и характеризует максимальное количество энергии, которое выделяется в единицу времени, а также максимальное количество ресинтезируемых АТФ в единицу времени.
В свою очередь, емкость энергетических систем, ограничивает продолжительность работы мышц.
 Источники энергии мышц
Источники энергии мышцРасщепление креатинфосфата
Молекула КрФ (креатинфосфат) восстанавливает АДФ (аденезиндифосфат) до АТФ (аденозинтрифосфат), путем перехода фосфатной группы от КрФ к АДФ с образованием креатина и АТФ:
- АДФ + КрФ ⇆ АТФ + креатин
Таким образом, креатинфосфат отвечает за постоянный уровень концентрации АТФ, за счет обратимой реакции перефосфорилирования.
В основном весь креатинфосфат содержится в скелетных мышцах, сердце, мозге и нервных тканях.
Образования свободного креатина стимулирует анаэробный гликолиз в медленных мышечных волокон (ММВ), и аэробный гликолиз в быстрых мышечных волокон (БМВ).
Для бодибилдинга, важную роль будет играть то, что полное восстановление запасов креатинфосфата происходит в течение 3-5 минут, и это возможно только после прекращения интенсивного силового тренинга, то есть в перерывах между подходами. Например, если бы, КрФ восстанавливал свой уровень во время силового упражнения, мы бы могли выполнять подход очень и очень долго, но этого не происходит, из-за ограниченного количества креатинфосфата в мышцах и его способа восстановления.
В отсутствии двигательной активности, то есть когда мышцы находятся в спокойном состоянии, концентрация КрФ в 3-8 раз превышает концентрацию АТФ, благодаря чему во время коротких и интенсивных периодов мышечной активности, креатинфосфат успешно справляется с нехваткой АТФ для выполнения мышечных сокращений.
Силовая нагрузка, выполняемая в тренажерном зале, на первых 5-10 секундах интенсивного тренинга затрачивает креатинфосфат, потом подключаться другой путь восстановления АТФ, анаэробный гликолиз.
 Взрывная сила и расход креатина в мышцах
Взрывная сила и расход креатина в мышцахТренировка фосфатной системы стоит на первостепенном месте у всех атлетов, которые в своей подготовке используют упражнения взрывного характера, начиная от быстрой пробежки, на короткие дистанции, заканчивая выполнением силового подхода в тренажерном зале на 2-4 повторения. К таким атлетам, прежде всего относят: спринтеров, футболистов, двоеборцев, борцов, боксеров, метателей дисков/ядер/копья, прыгунов в высоту/длину, боксеры, теннисисты, другими словами все спортсмены, которым на соревнованиям необходимо в тот или иной момент показать высокую физическую работоспособность в короткий промежуток времени.
Фосфагенная система считается очень быстрым, мощным источником восполнения энергии (в единицу времени мощность КрФ в 3 раза превышает мощность гликолиза, ив 4-10 раз окисления!), однако, емкость не велика, именно поэтому при очень интенсивной физической работы, атлет может продолжать усилия не более 5-6 секунд.
Когда запасы креатинфосфата заканчиваются, в саркоплазме запускаются процессы гликолиза.
Анаэробный гликолиз
Примерно на 20-30 секунде выполнения силового упражнения, запасы креатинфосфата исчерпываются, и для снабжения организма энергией, подключается в работу анаэробный гликолиз, то есть происходит расщепление углеводов (глюкозы) в отсутствии кислорода, до лактата (молочной кислоты) :
- С6Н1206 (глюкоза) + 2 Н3Р04 + 2 АДФ = 2 С3Н6О3 (лактат) + 2 АТФ (энергия) + 2 Н2O
В связи с тем, что в анаэробном гликолизе, в качестве побочного продукта выступает молочная кислота, которая очень медленно вымывается, то с течением времени ее все больше и больше накапливается в мышцах при их сокращении, по итогу, наши дальнейшие движения мышц становятся затруднительными, ощущается чувство жжение, в конце концов, сокращения прекращаются.
Молочная кислота выступает в роли блокатора, которая нарушает сигнал, от мотонейронов к мышцам.
При интенсивных тренировках, лактат часто используется организмом, в качестве источника энергии и сырья для производства глюкозы.
 Расход мышечного гликогена в тренажерном зале
Расход мышечного гликогена в тренажерном залеМолочная кислота нейтрализуется в мышцах практически полностью (на 95%), спустя 1 ч 15 минут. Для того чтобы ускорить этот процесс, мы рекомендуем использовать активный отдых и заминку после тренировки, так как 75% образуемой молочной кислоты, во время интенсивной нагрузки переходят из быстрых мышечных волокон в медленные, которые используются ими в качестве источника энергии, а как мы знаем, активный отдых это работа медленных мышечных волокон, именно поэтому, пассивных отдых уступает активному.
Аэробный гликолиз
На 80-90 секунде работы мышц, запускаются процессы аэробного гликолиза в митохондриях, для протекания которого необходим кислород.
На промежуточном этапе аэробного гликолиза образуется две молекулы пирувата, проникнув в митохондрии они окисляются благодаря циклу Кребса, до углекислого газа (CO2 ) и воды (h3O), с образованием 38 молекул АТФ.
- C6h22O6 + 6O2 + 38АДФ + 38h4PO4 = 6CO2 + 44h3O + 38АТФ
Таким образом, аэробное окисление глюкозы гораздо эффективнее, чем анаэробное, однако анаэробный гликолиз, это единственный способ снабжения организма энергией в условиях кислородного голодания. В силу большой эффективности аэробного гликолиза, по отношению к анаэробному (в 19 раз!), в тканях, при поступлении кислорода, анаэробный путь поступления энергии блокируется (эффект Пастера).
Виды типичных аэробных упражнений:
- Плавание
- Бег на средние и длинные дистанции
- Скакалка
- Быстрая ходьба
- Лыжи
- Велосипед
При этом помните, что в начале выполнения упражнения, энергообеспечение организма происходит за счет анаэробного гликолиза, но спустя 1-2 минуты, при сохранении активных движений, энергообеспечение уже происходит благодаря процессам аэробного окисления глюкозы. Конечно, если работа носит взрывной короткий характер, то первые 5-10 секунд энергообеспечение происходит за счет расщепление КрФ, а потом уже при снижении интенсивности, последовательно идет анаэробный гликолиз, и через 1-2 минуты аэробный гликолиз.
У не тренированных спортсменов, молочной кислоты во время физической нагрузки образуется очень много, что говорит о слабой аэробной системы, в свою очередь тренированные атлеты, могут похвастаться аэробными способностями, то есть высокой способностью усваивать организмом кислород.
 Бег и аэробный гликолиз
Бег и аэробный гликолизНапример, не подготовленные атлеты, начинают чувствовать достаточно быстро при беге/езде на велосипеде/лыжах болезненность в мышцах, в виде жжения, молочная кислота у них очень быстро образуется, при недостатке кислорода, таким образом, происходит нарушение кислотно-щелочного равновесия в мышцах (ацидоз), то есть их закисление.
При умеренной, продолжительной физической нагрузки, снабжения организма энергией происходит за счет использования мышечного гликогена, в течение следующих 30 минут, вклад глюкозы в энергообеспечение становиться все меньше и меньше, в конце концов, организм перестраивается на использование в качестве энергии жирных кислот.
Чем интенсивнее и меньше по продолжительности нагрузка, тем больше вклад анаэробного процесса образования АТФ, и наоборот, если нагрузка продолжительная, средней интенсивности, тем больше преобладают процессы аэробного синтеза АТФ.
Аэробное окисление углеводов и жиров
Когда ткани организма в достаточной мере снабжаться кислородом, на помощь в энергообеспечении приходят углеводы и липиды, являясь лучшим поставщиком энергии в аэробных условиях, то есть, кровь переносит к работающим мышцам жирные кислоты, глюкозу, они в свою очередь в митохондриях окисляются с выделением энергии.
На примере, пальмитиновой кислоты (жирная кислота), уравнение будет выглядеть так:
- Ch4(Ch3)14COOH + 23 O2 +129 АДФ = 6CO2 + 146 h3O + 129 АТФ
На примере глюкозы (углевода):
- C6h22O6 + O2 = CO2 + h3O + Q
Емкость окислительной системы наибольшая и превосходит во много раз все остальные, например, в 20 раз выше емкости гликолиза, именно поэтому, во время монотонной, продолжительной работы, в течении 1-2 часа, работу выполняет именно окислительная энергосистема.
Для обеспечения энергосистемы окисления, используются углеводы и жиры (липиды), причем, чем интенсивнее человек тренируется, тем больше расходуются углеводы, и меньше жиры.
 Марафонское плавание — как лучший способ сжигания жира
Марафонское плавание — как лучший способ сжигания жираМПК (максимальное потребление кислорода)
Условно, для того, охарактеризовать степень интенсивности выполняемой работы, ввели термин, МПК (максимальное потребление кислорода), который уникален у всех. Так, например, при выполнении легкой аэробной работы (легкий бег) используются жиры, в этом случае МПК не превышает 50%, при достижении МПК 70% и выше, начинают расходоваться уже углеводы, то есть при выполнении более тяжелой, интенсивной аэробной работы.
