гемолитическая, обтурационная, печеночно-клеточная. Желтуха новорожденных.
Первая
реакция катаболизма гема происходит
при участии NADPH-зависимого ферментативного
комплекса гемоксигеназы. Ферментная
сисгема локализована в мембране ЭР, в
области электронтранспортных цепей
микросомального окисления. Фермент
катализирует расщепление связи между
двумя пиррольными кольцами, содержащих
винильные остатки, — таким образом,
раскрывается структура кольца В
ходе реакции образуются линейный
тетрапир-рол — биливердин(пигмент
жёлтого цвета) и монооксид углерода
(СО), который получается из углерода
метениловой группы. Гем индуцирует
транскрипцию гена гемоксигеназы,
абсолютно специфичной по отношению к
тему. Ионы железа, освободившиеся при
распаде гема, могут быть использованы
для синтеза новых молекул гемоглобина
или для синтеза других железосодержащих
белков. Биливердин восстанавливается
до билирубина NADPH-зависимым
ферментом
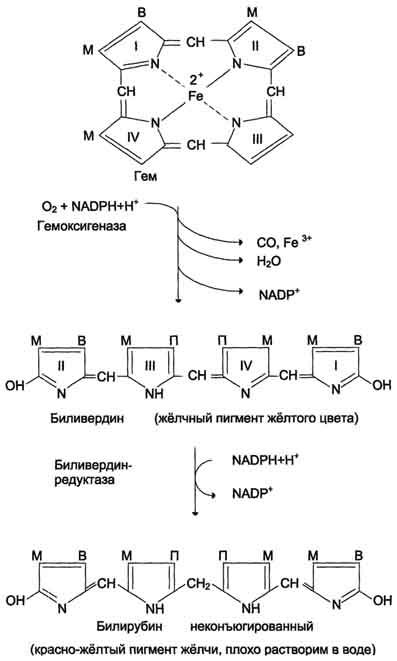
Билирубин, образованный в клетках РЭС (селезёнки и костного мозга), плохо растворим в воде, по крови транспортируется в комплексе с белком плазмы крови альбумином. Эту форму билирубина называют неконъюгированным билирубином. Каждая молекула альбумина связывает (или даже 3) молекулы билирубина, одна из которых связана с белком более прочно (более высокое сродство), чем другие. При сдвиге рН крови в кислую сторону (повышение концентрации кетоновых тел, лактата) изменяются заряд, конформация альбумина, снижается сродство к билирубину. Поэтому билирубин, связанный с альбумином непрочно, может вытесняться из центров связывания и образовывать комплексы с коллагеном межклеточного матрикса и липидами мембран. Ряд лекарственных соединений конкурирует с билирубином за высокоаффинный, имеющий высокое сродство центр альбумина.
Поглощение билирубина паренхиматозными клетками печени. Комплекс «альбумин-билирубин», доставляемый с током крови в печеНb, на поверхности плазматической мембраны гепатоцита диссоциирует. Высвобожденный билирубин образует временный комплекс с липидами плазматической мембраны. Облегчённая диффузия билирубина в гепатоциты осуществляется двумя типами белков-переносчиков: лигандина (он транспортирует основное количество билирубина) и протеина Z. Активность поглощения билирубина гепатоцитом зависит от скорости его метаболизма в клетке. Лигандин и протеин Z обнаружены также в клетках почек и кишечника, поэтому при недостаточности функции печени они способны компенсировать ослабление процессов детоксикации в этом органе.
Конъюгация билирубина в гладком ЭР. В гладком ЭР гепатоцитов к билирубину присоединяются (реакция конъюгации) полярные группы, главным образом от глюкуроновой кислоты.Билирубин имеет 2 карбоксильные группы, поэтому может соединяться с 2 молекулами глюкуроновой кислоты, образуя хорошо растворимый в воде конъюгат — диглюкуронид билирубина (конъюгированный, или прямой, билирубин).

Донором глюкуроновой кислоты служит УДФ-глюкуронат. Специфические ферменты, УДФ-глюкуронилтрансферазы (уридиндифосфоглюкуронилтрансферазы) катализируют образование моно- и диглюкуронидов билирубина.

Секреция
конъюгированного билирубина в жёлчь идёт по механизму активного транспорта,
т.е. против градиента концентрации.
Активный транспорт является, вероятно,
ско-рость-лимитирующей стадией всего
процесса метаболизма билирубина в
печени. В норме диглюкуронид билирубина
— главная форма экскреции билирубина в
жёлчь, однако не исключается присутствие
небольшого количества моноглюкуронида.
Транспорт конъюгирован-ного билирубина
из печени в жёлчь активируется теми же
лекарствами, которые способны индуцировать
конъюгацию билирубина. Таким образом,
можно сказать, что скорость конъюгации
билирубина и активный транспорт
билирубинглюкуронида из гепатоцитов
в жёлчь строго взаимосвязаны
В
кишечнике поступившие билирубинглюкурониды
гидролизуются специфическими
бактериальными ерментами β-глюкуронидазами,
которые гидролизуют связь между
билирубином и остатком глюкуроновой
кислоты. Освободившийся в ходе этой
реакции билирубин под действием кишечной
микрофлоры восстанавливается с
образованием группы бесцветных
тет-рапиррольных соединений –
Причинами гипербилирубинемии могут быть увеличение образования билирубина, превышающее способность печени экскретировать его, или повреждение печени, приводящее к нарушению секреции билирубина в жёлчь в нормальных количествах. Гипербилирубинемию отмечают также при закупорке желчевыводящих протоков печени. Во всех случаях содержание билирубина в крови повышается. При достижении определённой концентрации он диффундирует в ткани, окрашивая их в жёлтый цвет. Пожелтение тканей из-за отложения в них билирубина называют желтухой. Клинически желтуха может не проявляться до тех пор, пока концентрация билирубина в плазме крови не превысит верхний предел нормы более чем в 2,5 раза, т.е. не станет выше 50 мкмоль/л.
Гемолитическая (надпечёночная) желтуха. Известно, что способность печени образовывать глюкурониды и выделять их в жёлчь в 3-4 раза превышает их образование в физиологических условиях. Гемолитическая (надпечёночная) желтуха — результат интенсивного гемолиза эритроцитов. Она обусловлена чрезмерным образованием билирубина, превышающим способность печени к его выведению. Гемолитическая желтуха развивается при исчерпании резервных возможностей печени. Основная причина надпечёночной желтухи — наследственные или приобретённые гемолитические анемии. При гемолитических анемиях, вызванных сепсисом, лучевой болезнью, дефицитом глюкозо-6-фосфатдегидрогеназы эритроцитов, та-лассемией, переливанием несовместимых групп крови, отравлением сульфаниламидами, количество освобождающегося из эритроцитов гемоглобина за сутки может доходить до 45 г (при норме 6,25 г), что значительно увеличивает образование билирубина. Гипербилирубинемия у больных гемолитической желтухой обусловлена значительным повышением (103 — 171 мкмоль/л) в крови концентрации альбуминсвязанного не-конъюгированного билирубина (непрямой билирубин). Образование в печени и поступление в кишечник больших количеств билирубинглюкуронидов (прямой билирубин) ведёт к усиленному образованию и выделению с калом и мочой уробилиногенов и более интенсивной их окраски. Один из главных признаков гемолитической желтухи — повышение содержания в крови не-конъюгированиого (непрямого) билирубина. Это позволяет легко отличить её от механической (подпечёночной) и печёночно-клеточной (печёночной) желтух. Неконъюгированный билирубин токсичен. Гидрофобный, липофильный неконъюгирован-ный билирубин, легко растворяясь в липидах мембраны и проникая вследствие этого в митохондрии, разобщает в них дыхание и окислительное фосфорилирование, нарушает синтез белка, поток ионов калия через мембрану клетки и органелл. Это отрицательно сказывается а состоянии ЦНС, вызывая у больных ряд характерных неврологических симптомов.
Желтуха новорождённых. Частая разновидность гемолитической желтухи новорождённых — «физиологическая желтуха», наблюдающаяся в первые дни жизни ребёнка. Причиной повышения концентрации непрямого билирубина в крови служит ускоренный гемолиз и недостаточность функции белков и ферментов печени, ответственных за поглощение, конъюгацию и секрецию прямого билирубина. У новорождённых не только снижена активность УДФ-глюкуронилтрансферазы, но и, по-видимому, недостаточно активно происходит синтез второго субстрата реакции конъюгации УДФ-глюкуроната. Известно, что УДФ-глюкуронилтрансфераза — индуцируемый фермент. Одно из неприятных осложнений «физиологической желтухи» — билирубиновая энцефалопатия. Когда концентрация неконъюгированного билирубина превышает 340 мкмоль/л, он проходит через гематоэнцефалический барьер головного мозга и вызывает его поражение.
Печёночно-клеточная (печёночная) желтуха. Печёночно-клеточная (печёночная) желтуха обусловлена повреждением гепатоцитов и жёлчных капилляров, например, при острых вирусных инфекциях, хроническом и токсических гепатитах.Причина повышения концентрации билирубина в крови — поражение и некроз части печёночных клеток. Происходит задержка билирубина в печени, чему способствует резкое ослабление метаболических процессов в поражённых гепатоцитах, которые теряют способность нормально выполнять различные биохимические и физиологические процессы, в частности переводить конъюгированный (прямой) билирубин из клеток в жёлчь против градиента концентрации. Для печёночно-клеточной желтухи характерно то, что вместо преобладающих в норме диглюкуронидов билирубина в поражённой печёночной клетке образуются главным образом моноглюкурониды В результате деструкции печёночной паренхимы образующийся прямой билирубин частично попадает в большой круг кровообращения, что ведёт к желтухе. Экскреция жёлчи также нарушена. Билирубина в кишечник попадает меньше, чем в норме. При печёночно-клеточной желтухе повышается концентрация в крови как общего билирубина, так и обеих его фракций — неконъюгированного (непрямого) и конъюгированного (прямого). Так как в кишечник поступает меньше билирубинглюкуронида, то и количество образующегося уробилиногена также снижено. Поэтому кал гипохоличный, т.е. менее окрашенный. Моча, наоборот, имеет более интенсивную окраску за счёт присутствия там не только уробилинов, но и конъюгированного билирубина, который хорошо растворим в воде и экскретируется с мочой.
Механическая, или обтурационная (подпечёночная) желтуха. Механическая, или обтурационная (подпечёночная), желтуха развивается при нарушении желчеотделения в двенадцатиперстную кишку. Это может быть вызвано закупоркой жёлчных протоков, например при желчнокаменной болезни, опухолью поджелудочной железы, жёлчного пузыря, печени, двенадцатиперстной кишки, хроническим воспалением поджелудочной железы или послеоперационным сужением общего жёлчного протока При полной закупорке общего жёлчного протока конъюгированный билирубин в составе желчи не поступает в кишечник, хотя гепатоциты продолжают его вырабатывать. Поскольку билирубин в кишечник не попадает, продуктов его катаболизма уробилиногенов в моче и кале нет. Кал обесцвечен. Так как нормальные пути экскреции билирубина заблокированы, происходит его утечка в кровь, поэтому в крови больных повышена концентрация конъюгированного билирубина. Растворимый билирубин экскретируется с мочой, придавая ей насыщенный оранжево-коричневый цвет.
|
|
Продолжительность жизни эритроцитов составляет 120 дней, затем они разрушаются и освобождается гемоглобин. Главными органами, в которых происходят разрушение эритроцитов и распад гемоглобина, являются печень, селезенка и костный мозг, хотя в принципе оба процесса могут происходить и в клетках других органов. Распад гемоглобина в печени начинается с разрыва α-метиновой связи между I и II кольцами порфиринового кольца. Этот процесс катализируется НАДФ-содержащей ок-сидазой и приводит к образованию зеленого пигмента вердоглобина (холеглобина): В приведенных структурных формулах здесь и далее в желчных пигментах М – метильная СН3-группа, В – (—СН=СН2) – винильная группа и П – (—СН2—СН2—СООН) – остаток пропионовой кислоты. Как видно из приведенных формул, в молекуле вердоглобина еще сохраняются атом железа и белковый компонент. Имеются экспериментальные доказательства, что в этом окислительном превращении гемоглобина принимают участие витамин С, ионы Fe2+и другие кофакторы. Дальнейший распад вердоглобина, вероятнее всего, происходит спонтанно с освобождением железа, белка-глобина и образованием одного из желчных пигментов – биливердина. Спонтанный распад сопровождается перераспределением двойных связей и атомов водорода в пиррольных кольцах и метиновых мостиках. Образовавшийся биливердин ферментативным путем восстанавливается в печени в билирубин, являющийся основным желчным пигментом у человека и плотоядных животных: Основное место образования билирубина – печень, селезенка и, по-видимому, эритроциты (при распаде их иногда разрывается одна из метиновых связей в протопорфирине). Образовавшийся во всех этих клетках билирубин поступает в печень, откуда вместе с желчью попадает в желчный пузырь (см. главу 16). Билирубин, образовавшийся в клетках системы макрофагов, называется свободным, или непрямым, билирубином, поскольку вследствие плохой растворимости в воде он легко адсорбируется на белках плазмы крови и для его определения в крови необходимо предварительное осаждение белков спиртом. После этого билирубин вступает во взаимодействие с диазореактивом Эрлиха. В крови взрослого здорового человека содержится относительно постоянное количество общего билирубина – от 4 до 26 мкмоль/л, в среднем 15 мкмоль/л. Около 75% этого количества приходится на долю непрямого билирубина. Повышение его концентрации в крови до 35 мкмоль/л приводит к желтухе. Более высокий уровень билирубина в крови вызывает явления тяжелого отравления. Непрямой билирубин, поступая с током крови в печень, подвергается обезвреживанию путем связывания с глюку-роновой кислотой. В этом процессе принимают участие особый фермент УДФ-глюкуронилтрансфераза и УДФ-глюкуроновая кислота, являющаяся донором глюкуроновой кислоты. При этом к билирубину присоединяются 2 остатка глюкуроновой кислоты с образованием сравнительно индифферентного комплекса – билирубин-диглюкуронида, хорошо растворимого в воде и дающего прямую реакцию с диазореактивом. В желчи всегда присутствует прямой билирубин. В крови количество прямого и непрямого билирубина, а также соотношение между ними резко меняются при поражениях печени, селезенки, костного мозга, болезнях крови и т.д., поэтому определение содержания обеих форм билирубина в крови имеет существенное значение при дифференциальной диагностике различных форм желтухи. При желчнокаменной болезни в составе желчных камней наряду с основным их компонентом – холестерином всегда обнаруживается непрямой билирубин. Вследствие плохой растворимости в воде он выпадает в осадок в желчном пузыре в виде билирубината кальция, участвующего в формировании камней. Дальнейшая судьба желчных пигментов, точнее билирубина, связана с их превращениями в кишечнике под действием бактерий. Сначала глю-куроновая кислота отщепляется от комплекса с билирубином и освободившийся билирубин подвергается восстановлению в стеркобилиноген, который выводится из кишечника. В сутки человек выделяет около 300 мг стеркобилиногена. Последний легко окисляется под действием света и воздуха в стеркобилин. Механизм бактериальных превращений билирубина до стеркобилина до конца еще не расшифрован. Имеются данные, что промежуточными продуктами восстановления являются последовательно мезобилирубин и мезобилиноген (уробилиноген). После всасывания небольшая часть мезобилиногена поступает через воротную вену в печень, где подвергается разрушению с образованием моно- и дипиррольных соединений. Кроме того, очень небольшая часть стеркобилиногена после всасывания через систему геморроидальных вен попадает в большой круг кровообращения, минуя печень, и в таком виде выводится с мочой. Однако называть его уробилиногеном не совсем точно (см. главу 18). Суточное содержание стеркобилиногена в моче составляет около 4 мг, и, пожалуй, именно стеркобилиноген является нормальной органической составной частью мочи. Если с мочой выделяется повышенное содержание уробилиногена (точнее, мезобилиногена), то это является свидетельством недостаточности функции печени, например, при печеночной или гемолитической желтухе, когда печень частично теряет способность извлекать этот пигмент из крови воротной вены. Химически уробилиноген (мезо-билиноген) неидентичен стеркобилиногену (уробилиногену) мочи. Исчезновение стеркобилиногена (уробилиногена) из мочи при наличии билирубина и биливердина является свидетельством полного прекращения поступления желчи в кишечник. Такое состояние часто наблюдается при закупорке протока желчного пузыря (желчнокаменная болезнь) или общего желчного протока (желчнокаменная болезнь, раковые поражения поджелудочной железы и др.). Таким образом, количественный и качественный анализ желчных пигментов в моче может представлять большой клинический интерес. Предыдущая страница | Следующая страница СОДЕРЖАНИЕ |
42. Гемоглобин. Строение. Синтез и распад гемоглобина. Формы билирубина. Пути выведения билирубина и других желчных пигментов. Желтухи.
Гемоглобин входит в состав группы белков гемопротеины, которые сами являются подвидом хромопротеинов и подразделяются на неферментативные белки (гемоглобин, миоглобин) и ферменты (цитохромы, каталаза, пероксидаза). Небелковой частью их является гем – структура, включающая в себя порфириновое кольцо (состоящее из 4 пиррольных колец) и иона Fe2+. Железо связывается с порфириновым кольцом двумя координационными и двумя ковалентными связями.
Строение гемоглобина. Гемоглобин представляет собой белок, включающий 4 гемсодержащие белковые субъединицы. Между собой протомеры соединяются гидрофобными, ионными, водородными связями по принципу комплементарности. При этом они взаимодействуют не произвольно, а определенным участком — контактной поверхностью. Этот процесс высокоспецифичен, контакт происходит одновременно в десятках точек по принципу комплементарности. Взаимодействие осуществляют разноименно заряженные группы, гидрофобные участки, неровности на поверхности белка. Белковые субъединицы в нормальном гемоглобине могут быть представлены различными типами полипептидных цепей: α, β, γ, δ, ε, ξ (соответственно, греч. — альфа, бета, гамма, дельта, эпсилон, кси). В состав молекулы гемоглобина входят по две цепи двух разных типов. Гем состоит из иона двухвалентного железа и порфирина. В основе структуры порфиринов находится порфин. Порфин представляет собой четыре пиррольных кольца, связанных между собой метеновыми мостикамиГем соединяется с белковой субъединицей, во-первых, через остаток гистидина координационной связью железа, во-вторых, через гидрофобные связи пиррольных колец и гидрофобных аминокислот. Гем располагается как бы «в кармане» своей цепи и формируется гемсодержащий протомер.
Существует несколько нормальных вариантов гемоглобина:
HbР – примитивный гемоглобин, содержит 2ξ- и 2ε-цепи, встречается в эмбрионе между 7-12 неделями жизни,
HbF – фетальный гемоглобин, содержит 2α- и 2γ-цепи, появляется через 12 недель внутриутробного развития и является основным после 3 месяцев,
HbA – гемоглобин взрослых, доля составляет 98%, содержит 2α- и 2β-цепи, у плода появляется через 3 месяца жизни и к рождению составляет 80% всего гемоглобина,
HbA2 – гемоглобин взрослых, доля составляет 2%, содержит 2α- и 2δ-цепи,
HbO2 – оксигемоглобин, образуется при связывании кислорода в легких, в легочных венах его 94-98% от всего количества гемоглобина,
HbCO2 – карбогемоглобин, образуется при связывании углекислого газа в тканях, в венозной крови составляет 15-20% от всего количества гемоглобина.
Патологические формы гемоглобина
HbS – гемоглобин серповидно-клеточной анемии.
MetHb – метгемоглобин, форма гемоглобина, включающая трехвалентный ион железа вместо двухвалентного. Такая форма обычно образуется спонтанно, в этом случае ферментативных мощностей клетки хватает на его восстановление. При использовании сульфаниламидов, употреблении нитрита натрия и нитратов пищевых продуктов, при недостаточности аскорбиновой кислоты ускоряется переход Fe2+ в Fe3+. Образующийся metHb не способен связывать кислород и возникает гипоксия тканей. Для восстановления ионов железа в клинике используют аскорбиновую кислоту и метиленовую синь.
Hb-CO – карбоксигемоглобин, образуется при наличии СО (угарный газ) во вдыхаемом воздухе. Он постоянно присутствует в крови в малых концентрациях, но его доля может колебаться от условий и образа жизни. Угарный газ является активным ингибитором гем-содержащих ферментов, в частности, цитохромоксидазы 4-го комплекса дыхательной цепи.
HbA1С
– гликозилированный гемоглобин.
Концентрация его нарастает при хронической
гипергликемии и является хорошим
скрининговым показателем уровня глюкозы
крови за длительный период времени.
Биосинтез гема. Гем является небелковой частью многих гемопротеинов:
гемоглобин (до 85% общего количества гема организма), локализованный в эритроцитах и клетках костного мозга,
миоглобин скелетных мышц и миокарда (17%),
цитохромы дыхательной цепи и P450, цитохромоксидаза, гомогентизатоксидаза, пероксидаза, миелопероксидаза, каталаза, тиреопероксидаза и т.д. – менее 1%. Синтез гема в основном идет в предшественниках эритроцитов, клетках печени, почек, слизистой кишечника, и в остальных тканях. Первая реакция синтеза с участием δ-аминолевулинат-синтазы происходит в митохондриях. Следующая реакция при участии аминолевулинатдегидратазы (порфобилиноген-синтазы) протекает в цитозоле.Гем синтезируется во всех тканях, но с наибольшей скоростью в костном мозге и печени. В костном мозге гем необходим для синтеза гемоглобина в ретикулоцитах, в гепатоцитах — для образования цитохрома Р450.
Первая
реакция синтеза гема — образование
5-аминолевулиновой кислоты из глицина
и сук-цинил-КоА идёт в матриксе митохондрий,
где в ЦТК образуется один из субстратов
этой реакции — сукцинил-КоА. Эту реакцию
катализирует пиридоксальзависимый
фермент аминолевулинатсинтаза. Из
митохондрий 5-аминолевулиновая кислота
поступает в цитоплазму. В цитоплазме
проходят промежуточные этапы синтеза
гема: соединение 2 молекул 5-аминолевулиновой
кислоты молекулу порфобилиногена,
дезаминирование порфобилиногена с
образованием гидроксиметилбилана,
ферментативное превращение
гидроксиметилбилана в молекулу
уропор-фобилиногена III, декарбоксилирование
последнего с образованием копропорфириногена
III. Гидроксиметилбилан может также
нефермента-тивно превращаться в
уропорфириноген I, который декарбоксилируется
в копропорфирино-ген I. Из цитоплазмы
копропорфириноген III опять поступает
в митохондрии, где проходят заключительные
реакции синтеза гема. В результате двух
последовательных окислительных реакций
копропорфириноген III превращается в
протопорфириноген IX, а протопорфириноген
IX — в Протопорфирин IX. Фермент феррохела-таза,
присоединяя к протопорфирину IX
двухвалентное лентное железо, превращает
его в гем. Источником железа для синтеза
гема служит депонирующий железо белок
ферритин. Синтезированный гем, соединяясь
с α и β-полипепептидными цепями глобина,
образует гемоглобин. Гем регулирует
синтез глобина: при снижении скорости
синтеза гема синтез глобина в ретикулоцитах
тормозится. 
Регуляция синтеза гема
1. Скорость синтеза глобиновых цепей зависит от наличия гема, он ускоряет биосинтез «своих» белков.
2. Основным регуляторным ферментом синтеза гема является аминолевулинатсинтаза.
гем после взаимодействия с молекулой белка-репрессора формирует активный репрессорный комплекс, связывается с ДНК и подавляет транскрипцию, мРНК для фермента не образуется и синтез фермента прекращается. Также имеется отрицательный аллостерический эффект гема на фермент.
с другой стороны, достаточное количество ионов железа оказывает положительный эффект при синтезе молекулы аминолевулинатсинтазы. В клетке имеется особый железосвязывающий белок, который в отсутствии ионов железа обладает сродством к мРНК фермента и блокирует ее трансляцию в рибосоме, т.е. синтез белковой цепи. Ионы железа связываются с этим железосвязывающим белком, образуя с ним неактивный комплекс, что инициирует синтез фермента.
3. Положительным модулятором аминолевулинатсинтазы служит гипоксия тканей, которая в эритропоэтических тканях индуцирует синтез фермента.
4. В печени повышение активности аминолевулинатсинтазы вызывают соединения, усиливающие работу микросомальной системы окисления (жирорастворимые токсины, стероиды) – при этом возрастает потребление гема для образования цитохрома Р450, что снижает внутриклеточную концентрацию свободного гема. В результате происходит дерепрессия синтеза фермента.
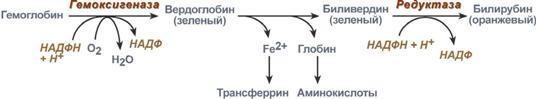
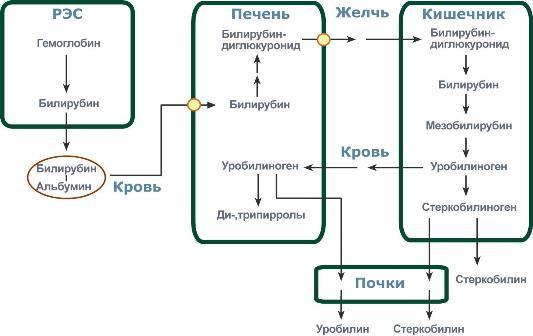
Распад. За сутки у человека распадается около 9 г гемопротеинов, в основном это гемоглобин эритроцитов. Эритроциты живут 90-120 дней, после чего лизируются в кровеносном русле или в селезенке. При разрушении эритроцитов в кровяном русле высвобождаемый гемоглобин образует комплекс с белком-переносчиком гаптоглобином (фракция α2-глобулинов крови) и переносится в клетки ретикуло-эндотелиальной системы (РЭС) селезенки (гл образом), печени и костного мозга.
Первая реакция катаболизма гема происходит при участии NADPH-зависимого ферментативного комплекса гемоксигеназы. Ферментная сисгема локализована в мембране ЭР, в области электронтранспортных цепей микросомального окисления. Фермент катализирует расщепление связи между двумя пиррольными кольцами, содержащих винильные остатки, — таким образом, раскрывается структура кольца. В ходе реакции образуются линейный тетрапир-рол — биливердин (пигмент жёлтого цвета) и монооксид углерода (СО), который получается из углерода метениловой группы. Гем индуцирует транскрипцию гена гемоксигеназы, абсолютно специфичной по отношению к тему.
Ионы железа, освободившиеся при распаде гема, могут быть использованы для синтеза новых молекул гемоглобина или для синтеза других железосодержащих белков. Биливердин восстанавливается до билирубина NADPH-зависимым ферментом биливердинредуктазой. Билирубин образуется не только при распаде гемоглобина, не также при катаболизме других гемсодержащю белков, таких как цитохромы и миоглобин. При распаде 1 г гемоглобина образуется 35 мг билирубина, а в сутки у взрослого человека — примерно 250-350 мг билирубина. Дальнейший метаболизм билирубина происходит в печени.
В
клетках РЭС гем в составе гемоглобина
окисляется молекулярным кислородом. В
реакциях последовательно происходит
разрыв метинового мостика между 1-м и
2-м пиррольными кольцами гема с их
восстановлением, отщеплением железа и
белковой части и образованием оранжевого
пигмента билирубина. Билирубин – токсичное, жирорастворимое вещество,
способное нарушать окислительное
фосфорилирование в клетках. Особенно
чувствительны к нему клетки нервной
ткани. Из клеток ретикуло-эндотелиальной
системы билирубин попадает в кровь.
Здесь он находится в комплексе с
альбумином плазмы, в гораздо меньшем
количестве – в комплексах с металлами,
аминокислотами, пептидами и другими
малыми молекулами. Образование таких
комплексов не позволяет выделяться
билирубину с мочой. Билирубин в комплексе
с альбумином называется свободный
(неконъюгированный) или непрямой
билирубин. Из сосудистого русла в
гепатоциты билирубин попадает с помощью
белка-переносчика (лигандина). В клетке
протекает реакция связывания билирубина
с УДФ-глюкуроновой кислотой, при этом
образуются моно- и диглюкурониды. Кроме
глюкуроновой кислоты, в реакцию могут
вступать сульфаты, фосфаты, глюкозиды.
Билирубин-глюкуронид получил название
связанный (конъюгированный) или прямой
билирубин. После образования
билирубин-глюкурониды АТФ-зависимым
переносчиком секретируются в желчные
протоки и далее в кишечник, где при
участии бактериальной β-глюкуронидазы
превращаются в свободный билирубин.
Одновременно некоторое количество
билирубин-глюкуронидов может попадать
(особенно у взрослых) из желчи в кровь
по межклеточным щелям. Таким образом,
в крови в норме одновременно существуют
две формы билирубина: свободный,
попадающий сюда из клеток РЭС (около
80% всего количества), и связанный,
попадающий из желчных протоков (до 20%). Превращение
в кишечнике.
В кишечнике билирубин подвергается
восстановлению под действием микрофлоры
до мезобилирубина и мезобилиногена
(уробилиногена). Часть последних
всасывается и с током крови вновь
попадает в печень, где окисляется до
ди- и трипирролов. При этом в здоровом
организме в общий круг кровообращения
и в мочу мезобилирубин и уробилиноген
не попадают, а полностью задерживаются
гепатоцитами. Оставшаяся в кишечнике
часть пигментов ферментами бактериальной
флоры толстого кишечника восстанавливается
до стеркобилиногена и выделяется из
организма, окрашивая кал. Незначительное
количество стеркобилиногена через
геморроидальные вены попадает в большой
круг кровообращения, отсюда в почки и
выделяется с мочой. На воздухе
стеркобилиноген и уробилиноген
превращаются, соответственно, в
стеркобилин и уробилин.
Ситуации,
при которых в крови накапливается
билирубин, в зависимости от причины
делятся на три вида:
1.Гемолитические – в результате гемолиза при избыточном превращении гемоглобина в билирубин,
2.Печеночно-клеточные – когда печень не в состоянии обезвредить билирубин,
3.Механические – если билирубин не может попасть из печени в кишечник из-за механического перекрытия желчевыводящих путей.
Накопление билирубина в крови свыше 43 мкмоль/л ведет к связыванию его эластическими волокнами кожи и конъюнктивы, что проявляется в виде желтухи. Так как свободный билирубин липофилен, то он легко накапливается в подкожном жире и нервной ткани. Последнее особенно опасно для детей, особенно для новорожденнных.
Гемолитическая или надпеченочная желтуха – ускоренное образование билирубина в результате усиления внутрисосудистого гемолиза. К данному типу желтух относятся гемолитические анемии различного происхождения: врожденный сфероцитоз, серповидно-клеточная анемия, дефицит глюкозо-6-фосфатдегидрогеназы, отравление сульфаниламидами, талассемии, сепсис, лучевая болезнь, несовместимость крови. В данном случае гипербилирубинемия развивается за счет фракции непрямого билирубина. Гепатоциты усиленно переводят непрямой билирубин в связанную форму, секретируют его в желчь, в результате в кале увеличивается содержание стеркобилина, интенсивно его окрашивая. В моче возрастает содержание уробилина, билирубин отсутствует.У новорожденных гемолитическая желтуха может развиться как симптом гемолитической болезни новорожденного.
Механическая или подпеченочная желтуха развивается вследствие снижения оттока желчи при непроходимости желчного протока – желчные камни, новообразования поджелудочной железы, гельминтозы. В результате застоя желчи происходит растяжение желчных капилляров, увеличивается проницаемость их стенок. Не имеющий оттока в кишечник прямой билирубин поступает в кровь, в результате развивается гипербилирубинемия. В тяжелых случаях, вследствие переполнения гепатоцитов прямым билирубином, реакция конъюгации с глюкуроновой кислотой может нарушаться, присоединяется печеночно-клеточная желтуха (см ниже). В результате в крови увеличивается концентрация непрямого билирубина. В моче резко увеличен уровень билирубина (цвет темного пива) и снижено количество уробилина, практически отсутствует стеркобилин кала (серовато-белое окрашивание).
Паренхиматозная (печеночно-клеточная) желтуха – причиной может быть нарушение на всех трех стадиях превращения билирубина в печени: извлечение билирубина из крови печеночными клетками, его конъюгирование и секреция в желчь. Наблюдается при вирусных и других формах гепатитов, циррозе и опухолях печени, жировой дистрофии печени, при отравлении токсическими гепатотропными веществами, при врожденных патологиях. Так как нарушаются все процессы превращения билирубина в печени, гипербилирубинемия развивается за счет обеих фракций, преимущественно прямого билирубина. Концентрация его возрастает из-за нарушения секреции в желчь и увеличения проницаемости мембран клеток печени. Количество непрямого билирубина возрастает за счет функциональной недостаточности гепатоцитов и/или снижения их количества. В моче определяется билирубин (цвет крепкого черного чая), умеренно увеличена концентрация уробилина, уровень стеркобилина кала в норме или снижен.
Гемолитическая болезнь новорожденного.Причины: Несовместимость крови матери и плода по группе или по резус-фактору. Накопление гидрофобной формы билирубина в подкожном жире обуславливает желтушность кожи. Однако реальную опасность представляет накопление билирубина в сером веществе нервной ткани и ядрах ствола с развитием «ядерной желтухи» (билирубиновая энцефалопатия).
Клиническая диагностика: Проявляется сонливостью, плохим сосанием, умственной отсталостью, ригидностью затылочных мышц, тоническими судорогами, тремором конечностей, изменением рефлексов с возможным развитием глухоты и параличей.
Физиологическая (транзиторная) желтуха новорожденных. Причины: относительное снижение активности УДФ-глюкуронилтрансферазы в первые дни жизни, связанное с повышенным распадом фетального гемоглобина, абсолютное снижение активности УДФ-глюкуронилтрансферазы в первые дни жизни, дефицит лигандина, слабая активность желчевыводящих путей. Клиническая диагностика: окрашивание кожи на 3-4 день после рождения, гемолиза и анемии нет. Симптомы исчезают спустя 1-2 недели после рождения.
Желтуха недоношенных.Причины: относительное снижение активности УДФ-глюкуронилтрансферазы в первые дни жизни, связанное с повышенным распадом фетального гемоглобина, абсолютное снижение активности УДФ-глюкуронилтрансферазы в первые дни жизни, дефицит лигандина, слабая активность желчевыводящих путей.Клиническая диагностика:окрашивание кожи, гемолиза и анемии нет. Исчезает спустя 3-4 недели после рождения.
— синтез и распад гема
Пигментный обмен — это совокупность сложных превращений окрашенных веществ тканей и биологических жидкостей. Большинство пигментов имеет белковое происхождение и являются дериватами порфириновых структур и ароматических соединений.
Одним из важнейших пигментов является гем, входящий в состав гемоглобина, миоглобина, цитохромов, каталазы. Вторую группу пигментов составляют предшественники гема — порфирины. Третья представлена продуктами распада гема — желчными пигментами. К четвертой группе относятся меланины — пигменты кожи, волос и роговицы глаза, являющиеся продуктом полимеризации диоксииндола, образующегося в процессе обмена тирозина.
Дефекты ферментов синтеза гема и метаболизма порфиринов подразделяются на первичные — наследственные энзимопатии, получившие название «порфирии», и вторичные — приобретенные, составляющие группу под названием «порфиринурии». Эти заболевания сопровождаются накоплением и выведением в избытке с мочой и калом различных производных порфиринов.
Основная часть желчных пигментов образуется при распаде гема и гемоглобина в клетках ретикулоэндотелиальной системы (РЭС) и представляет собой многоступенчатый процесс: при окислении гемоглобина образуется вердоглобин; после отщепления глобина и железа образуется биливердин, который далее восстанавливается до билирубина. Билирубин — желто-красный пигмент, представляющий собой линейный тетрапиррол, токсичное, жирорастворимое вещество, способное нарушать окислительное фосфорилирование в клетках, в первую очередь в нервной ткани. В крови билирубин либо находится в свободном состоянии, либо в комплексе с альбумином (частично в виде альбумин-фосфатидного комплекса), в меньшем количестве — в комплексах с металлами, аминокислотами, пептидами и другими малыми молекулами. Образование таких комплексов предотвращает выделение билирубина с мочой. Такая форма билирубина называется свободной (неконъюгированной, несвязанной, непрямой). Она не дает прямой реакции с диазореактивом Эрлиха.
Из сосудистого русла в гепатоциты билирубин попадает с помощью белка-переносчика (лигандина). В печени, при участии фермента УДФ‑глюкуронилтрансферазы, происходит реакция этерификации — взаимодействие OH‑группы глюкуроновой кислоты с карбоксильными группами билирубина и образование конъюгированного (связанного, прямого) билирубина, представляющего собой парное соединение с одним или двумя остатками глюкуроновой кислоты (моно- или диглюкуронид). Кроме глюкуроновой кислоты в реакцию могут вступать сульфаты, фосфаты, глюкозиды. В норме конъюгированный билирубин активно секретируется в желчные капилляры, где при участии β‑глюкуронидазы, вновь превращается в свободный билирубин и с током желчи попадает в тонкий кишечник. Здесь ферментами бактериальной флоры он восстанавливается до мезобилирубина и мезобилиногена (уробилиногена). Часть последних всасывается и с током крови вновь попадает в печень, где окисляется до ди‑ и трипирролов. При этом в здоровом организме в общий круг кровообращения и в мочу мезобилирубин и уробилиноген не попадают, а полностью задерживаются гепатоцитами. Невсосавшаяся часть пигментов ферментами бактериальной флоры толстого кишечника восстанавливается до стеркобилиногена и выделяется из организма, окрашивая кал. Незначительное количество стеркобилиногена через геморроидальные вены попадает в большой круг кровообращения, отсюда –– в почки и выделяется с мочой. На воздухе стеркобилиноген и уробилиноген превращаются, соответственно, в стеркобилин и уробилин.
В клинике состояние пигментного обмена определяют, оценивая:
- Количество промежуточных и конечных метаболитов синтеза порфириновых структур (порфобилиногена, δ‑аминолевулиновой кислоты, копропорфирина, уропорфирина) и активность δ‑аминолевулинат-дегидратазы.
- Уровень желчных пигментов — продуктов распада гемоглобина и других хромопротеидов (общего билирубина и его фракций).
Распад билирубина — Давление и всё о нём
За сутки у человека распадается около 9 г гемопротеинов, в основном это гемоглобин эритроцитов.
Эритроциты в норме живут 90-120 дней, после чего лизируются в клетках ретикулоэндотелиальной системы – макрофагах селезенки (главным образом), купферовских клетках печени и макрофагах костного мозга. При разрушении эритроцитов в кровеносном русле высвобождаемый гемоглобин образует комплекс с белком-переносчиком гаптоглобином (фракция α2-глобулинов крови) и также переносится в клетки РЭС селезенки, печени и костного мозга.
Синтез билирубина
В клетках РЭС гем в составе гемоглобина окисляется молекулярным кислородом. В реакциях последовательно происходит разрыв метинового мостика между 1-м и 2-м пиррольными кольцами гема с их восстановлением, отщеплением железа и белковой части и образованием оранжевого пигмента билирубина. Высвобождаемое железо может либо запасаться в клетке в комплексе с ферритином, либо выделяться наружу и связываться с трансферрином.

Реакции распада гемоглобина и образования билирубина
Билирубин – токсичное, жирорастворимое вещество, способное разобщать окислительное фосфорилирование в клетках. Особенно чувствительны к нему клетки нервной ткани.
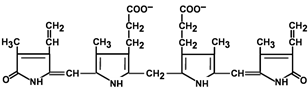
Строение билирубина
Выведение билирубина
Из клеток ретикуло-эндотелиальной системы билирубин попадает в кровь. Здесь он находится в комплексе с альбумином плазмы, в гораздо меньшем количестве – в комплексах с металлами, аминокислотами, пептидами и другими малыми молекулами. Образование таких комплексов не позволяет выделяться билирубину с мочой. Билирубин в комплексе с альбумином называется свободный (неконъюгированный) или непрямой билирубин.

Этапы метаболизма билирубина в организме
Из сосудистого русла в гепатоциты билирубин попадает с помощью белка-переносчика (транспортный белок органических анионов) или по механизму флип-флоп. Далее при участии цитозольного связывающего белка лигандина (Y-протеин) билирубин транспортируется в ЭПР, где протекает реакция связывания билирубина с УДФ-глюкуроновой кислотой, при этом образуются моно— и диглюкурониды. Кроме глюкуроновой кислоты, в реакцию конъюгации могут вступать сульфаты, фосфаты, глюкозиды.
Билирубин-глюкуронид получил название связанный (конъюгированный) или прямой билирубин.
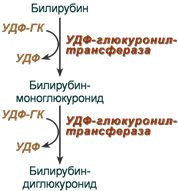 | 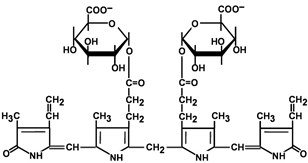 |
Реакции синтеза билирубин-диглюкуронида | Строение билирубин-диглюкуронида |
После образования билирубин-глюкурониды АТФ-зависимым переносчиком секретируются в желчные протоки и далее в кишечник, где при участии бактериальной β-глюкуронидазы превращаются в свободныйбилирубин. Одновременно, даже в норме (особенно у взрослых), некоторое количество билирубин-глюкуронидов может попадать из желчи в кровь по межклеточным щелям.
Таким образом, в плазме крови обычно присутствуют две формы билирубина: свободный (непрямой), попадающий сюда из клеток РЭС (80% и более всего количества), и связанный (прямой), попадающий из желчных протоков (не более 20%).
Термины «связанный«, «конъюгированный«, «свободный«, «несвязанный» отражают взаимодействие билирубина и глюкуроновой кислоты (но не билирубина и альбумина!).
Термины «прямой» и «непрямой» введены, исходя из возможности химической реакции билирубина с диазореактивом Эрлиха. Связанный билирубин реагирует с реактивом напрямую, без добавления дополнительных реагентов, т.к. является водорастворимым. Несвязанный (жирорастворимый) билирубин требует добавочных реактивов, реагирует не прямо.
Превращение в кишечнике
В кишечнике билирубин подвергается восстановлению под действием микрофлоры до мезобилирубина и мезобилиногена (уробилиногена). Часть уробилиногена всасывается и с кровью портальной вены попадает в печень, где либо распадается до моно-, ди- и трипирролов, либо окисляется до билирубина и снова экскретируется. При этом при здоровой печени в общий круг кровообращения и в мочу мезобилирубин и уробилиноген не попадают, а полностью задерживаются гепатоцитами.
Оставшаяся в кишечнике часть пигментов ферментами бактериальной флоры толстого кишечника восстанавливается до стеркобилиногена. Малая часть стеркобилиногена может всасываться и катаболизировать в печени, подобно уробилиногену. Также незначительное количество стеркобилиногена через геморроидальные вены попадает в большой круг кровообращения, отсюда в почки и выделяется с мочой (стеркобилин мочи). Основное количество стеркобилиногена достигает нижних отделов толстого кишечника и выделяется из организма.
В прямой кишке и на воздухе стеркобилиноген окисляется в стеркобилин, окрашивая кал. Аналогично уробилиноген, появляющийся в моче при патологии печени превращается в уробилин.
Очень часто стеркобилиноген, содержащийся в нормальной моче, называют уробилиногеном. И в клинической практике обычно не проводят различий между стеркобилиногеном и уробилиногеном мочи, их рассматривают как один пигмент – урохромы (уробилиноиды), что может создавать некоторую путаницу при оценке результатов анализа.
Вы можете спросить или оставить свое мнение.
Source: biokhimija.ru
Читайте также
ЛЕКЦИЯ Распад гема Желтухи Распад гема —
 ЛЕКЦИЯ «Распад гема» Желтухи
ЛЕКЦИЯ «Распад гема» Желтухи
 Распад гема — многостадийный процесс За сутки у человека распадается около 9 г гемопротеинов, в основном это гемоглобин эритроцитов. Эритроциты живут 90 -120 дней, после чего лизируются в кровеносном русле или в селезенке.
Распад гема — многостадийный процесс За сутки у человека распадается около 9 г гемопротеинов, в основном это гемоглобин эритроцитов. Эритроциты живут 90 -120 дней, после чего лизируются в кровеносном русле или в селезенке.
 4 основные этапа распада гема: I этап- внутри эритроцита старый гемоглобин подвергается частичной денатурации. II этап- катаболизм гема, освобождённого из любых гемовых белков, осуществляется в микросомальной фракции ретикуло – эндотелиальных клеток системой гемоксигеназы. III этап — превращения билирубина в печени : поглощение билирубина паренхиматозными клетками печени, коньюгация и секреция билирубина в желчь IV этап — метаболизм билирубина в кишечнике.
4 основные этапа распада гема: I этап- внутри эритроцита старый гемоглобин подвергается частичной денатурации. II этап- катаболизм гема, освобождённого из любых гемовых белков, осуществляется в микросомальной фракции ретикуло – эндотелиальных клеток системой гемоксигеназы. III этап — превращения билирубина в печени : поглощение билирубина паренхиматозными клетками печени, коньюгация и секреция билирубина в желчь IV этап — метаболизм билирубина в кишечнике.
 При разрушении эритроцитов в кровяном русле высвобождаемый гемоглобин образует комплекс с белком-переносчиком гаптоглобином (фракция α 2 -глобулинов крови) и переносится в клетки ретикулоэндотелиальной системы (РЭС) селезенки (главным образом), печени и костного мозга.
При разрушении эритроцитов в кровяном русле высвобождаемый гемоглобин образует комплекс с белком-переносчиком гаптоглобином (фракция α 2 -глобулинов крови) и переносится в клетки ретикулоэндотелиальной системы (РЭС) селезенки (главным образом), печени и костного мозга.
 Синтез билирубина В клетках РЭС гем в составе гемоглобина окисляется молекулярным кислородом. В реакциях последовательно происходит разрыв метинового мостика между 1 -м и 2 -м пиррольными кольцами гема с их восстановлением, отщеплением железа и белковой части и образованием оранжевого пигмента билирубина.
Синтез билирубина В клетках РЭС гем в составе гемоглобина окисляется молекулярным кислородом. В реакциях последовательно происходит разрыв метинового мостика между 1 -м и 2 -м пиррольными кольцами гема с их восстановлением, отщеплением железа и белковой части и образованием оранжевого пигмента билирубина.
 Билирубин – токсичное, жирорастворимое вещество, способное нарушать окислительное фосфорилирование в клетках. Особенно чувствительны к нему клетки нервной ткани.
Билирубин – токсичное, жирорастворимое вещество, способное нарушать окислительное фосфорилирование в клетках. Особенно чувствительны к нему клетки нервной ткани.
 Выведение билирубина Из клеток РЭС билирубин попадает в кровь. Здесь он находится в комплексе с альбумином плазмы, в гораздо меньшем количестве – в комплексах с металлами, аминокислотами, пептидами и другими малыми молекулами. Образование таких комплексов не позволяет выделяться билирубину с мочой. Билирубин в комплексе с альбумином называется свободный (неконъюгированный) или непрямой билирубин.
Выведение билирубина Из клеток РЭС билирубин попадает в кровь. Здесь он находится в комплексе с альбумином плазмы, в гораздо меньшем количестве – в комплексах с металлами, аминокислотами, пептидами и другими малыми молекулами. Образование таких комплексов не позволяет выделяться билирубину с мочой. Билирубин в комплексе с альбумином называется свободный (неконъюгированный) или непрямой билирубин.
. В клетке протекает") Из сосудистого русла в гепатоциты билирубин попадает с помощью белкапереносчика (лигандина). В клетке протекает реакция связывания билирубина с УДФ-глюкуроновой кислотой, при этом образуются моно- и диглюкурониды. Кроме глюкуроновой кислоты, в реакцию могут вступать сульфаты, фосфаты, глюкозиды. Билирубин-глюкуронид получил название связанный (конъюгированный) или прямой билирубин.
Из сосудистого русла в гепатоциты билирубин попадает с помощью белкапереносчика (лигандина). В клетке протекает реакция связывания билирубина с УДФ-глюкуроновой кислотой, при этом образуются моно- и диглюкурониды. Кроме глюкуроновой кислоты, в реакцию могут вступать сульфаты, фосфаты, глюкозиды. Билирубин-глюкуронид получил название связанный (конъюгированный) или прямой билирубин.
 Реакции синтеза билирубин-диглюкуронида
Реакции синтеза билирубин-диглюкуронида
 После образования билирубинглюкурониды АТФ-зависимым переносчиком секретируются в желчные протоки и далее в кишечник, где при участии бактериальной β-глюкуронидазы превращаются в свободный билирубин.
После образования билирубинглюкурониды АТФ-зависимым переносчиком секретируются в желчные протоки и далее в кишечник, где при участии бактериальной β-глюкуронидазы превращаются в свободный билирубин.
 из желчи в") • Одновременно некоторое количество билирубин-глюкуронидов может попадать (особенно у взрослых) из желчи в кровь по межклеточным щелям. • Таким образом, в крови в норме одновременно существуют две формы билирубина: свободный, попадающий сюда из клеток РЭС (около 80% всего количества), и связанный, попадающий из желчных протоков (до 20%).
• Одновременно некоторое количество билирубин-глюкуронидов может попадать (особенно у взрослых) из желчи в кровь по межклеточным щелям. • Таким образом, в крови в норме одновременно существуют две формы билирубина: свободный, попадающий сюда из клеток РЭС (около 80% всего количества), и связанный, попадающий из желчных протоков (до 20%).
 Превращение в кишечнике В кишечнике билирубин подвергается восстановлению под действием микрофлоры до мезобилирубина и мезобилиногена (уробилиногена). Часть последних всасывается и с током крови вновь попадает в печень, где окисляется до ди- и трипирролов. При этом в здоровом организме в общий круг кровообращения и в мочу мезобилирубин и уробилиноген не попадают, а полностью задерживаются гепатоцитами.
Превращение в кишечнике В кишечнике билирубин подвергается восстановлению под действием микрофлоры до мезобилирубина и мезобилиногена (уробилиногена). Часть последних всасывается и с током крови вновь попадает в печень, где окисляется до ди- и трипирролов. При этом в здоровом организме в общий круг кровообращения и в мочу мезобилирубин и уробилиноген не попадают, а полностью задерживаются гепатоцитами.
 Оставшаяся в кишечнике часть пигментов ферментами бактериальной флоры толстого кишечника восстанавливается до стеркобилиногена и выделяется из организма, окрашивая кал. Незначительное количество стеркобилиногена через геморроидальные вены попадает в большой круг кровообращения, отсюда в почки и выделяется с мочой. На воздухе стеркобилиноген и уробилиноген превращаются, соответственно, в стеркобилин и уробилин.
Оставшаяся в кишечнике часть пигментов ферментами бактериальной флоры толстого кишечника восстанавливается до стеркобилиногена и выделяется из организма, окрашивая кал. Незначительное количество стеркобилиногена через геморроидальные вены попадает в большой круг кровообращения, отсюда в почки и выделяется с мочой. На воздухе стеркобилиноген и уробилиноген превращаются, соответственно, в стеркобилин и уробилин.
 Этапы метаболизма билирубина в организме
Этапы метаболизма билирубина в организме
 Диагностическое значение определения билирубина плазмы крови Общий билирубин 1, 7 – 17 мкмоль/л 75% от общего билирубина непрямой билирубин (неконъюгированный) 25% — прямой, холебилирубин
Диагностическое значение определения билирубина плазмы крови Общий билирубин 1, 7 – 17 мкмоль/л 75% от общего билирубина непрямой билирубин (неконъюгированный) 25% — прямой, холебилирубин
 В клинике конъюгированный билирубин называют прямым, аом что он потому водорастворим и может быстро взаимодействовать с диазореагентом, образуя соединение розового цвета, — это прямая реакция Ван дер Берга. Неконъюгированный билирубингидрофобен, поэтому в плазме содержится в комплексе с альбумином и не реагирует с диазореактивом до тех пор, пока не добавлен органический растворитель, н-р этанол, который осаждает альбумин.
В клинике конъюгированный билирубин называют прямым, аом что он потому водорастворим и может быстро взаимодействовать с диазореагентом, образуя соединение розового цвета, — это прямая реакция Ван дер Берга. Неконъюгированный билирубингидрофобен, поэтому в плазме содержится в комплексе с альбумином и не реагирует с диазореактивом до тех пор, пока не добавлен органический растворитель, н-р этанол, который осаждает альбумин.
 2. Повреждение печени - угнетение") Причины гипербилирубинемии : 1. Увеличение образования билирубина (гемолитическая желтуха) 2. Повреждение печени — угнетение конъюгационных или выделительных механизмов печени (паренхиматозная желтуха) 3. Нарушение нормального пассажа желчи (механическая желтуха)
Причины гипербилирубинемии : 1. Увеличение образования билирубина (гемолитическая желтуха) 2. Повреждение печени — угнетение конъюгационных или выделительных механизмов печени (паренхиматозная желтуха) 3. Нарушение нормального пассажа желчи (механическая желтуха)
 Причины основных нарушений обмена билирубина
Причины основных нарушений обмена билирубина
 Накопление билирубина в крови свыше 43 мкмоль/л ведет к связыванию его эластическими волокнами кожи и конъюнктивы, что проявляется в виде желтухи. Так как свободный билирубин липофилен, то он легко накапливается в подкожном жире и нервной ткани.
Накопление билирубина в крови свыше 43 мкмоль/л ведет к связыванию его эластическими волокнами кожи и конъюнктивы, что проявляется в виде желтухи. Так как свободный билирубин липофилен, то он легко накапливается в подкожном жире и нервной ткани.
 Гемолитическая или надпеченочная желтуха – ускоренное образование билирубина в результате усиления внутрисосудистого гемолиза.
Гемолитическая или надпеченочная желтуха – ускоренное образование билирубина в результате усиления внутрисосудистого гемолиза.
 К данному типу желтух относятся гемолитические анемии различного происхождения: — врожденный сфероцитоз, — серповидно-клеточная анемия, — дефицит глюкозо-6 фосфатдегидрогеназы, отравление сульфаниламидами, — талассемии, сепсис, лучевая болезнь, — несовместимость крови. В данном случае гипербилирубинемия развивается за счет фракции непрямого билирубина.
К данному типу желтух относятся гемолитические анемии различного происхождения: — врожденный сфероцитоз, — серповидно-клеточная анемия, — дефицит глюкозо-6 фосфатдегидрогеназы, отравление сульфаниламидами, — талассемии, сепсис, лучевая болезнь, — несовместимость крови. В данном случае гипербилирубинемия развивается за счет фракции непрямого билирубина.
 • Гепатоциты усиленно переводят непрямой билирубин в связанную форму, секретируют его в желчь, в результате в кале увеличивается содержание стеркобилина, интенсивно его окрашивая. В моче возрастает содержание уробилина, билирубин отсутствует. • У новорожденных гемолитическая желтуха может развиться как симптом гемолитической болезни новорожденного.
• Гепатоциты усиленно переводят непрямой билирубин в связанную форму, секретируют его в желчь, в результате в кале увеличивается содержание стеркобилина, интенсивно его окрашивая. В моче возрастает содержание уробилина, билирубин отсутствует. • У новорожденных гемолитическая желтуха может развиться как симптом гемолитической болезни новорожденного.
 Один из главных признаков гемолитической желтухи – повышение в крови непрямого билирубина, который токсичен, легко растворяется в липидах мембраны и проникает вследствие этого в митохондрии, разобщая в них дыхание и окислительное фосфорилирование, нарушает синтез белков, поток ионов калия через мембрану клетки и органелл. Это отрицательно влияет на ЦНС.
Один из главных признаков гемолитической желтухи – повышение в крови непрямого билирубина, который токсичен, легко растворяется в липидах мембраны и проникает вследствие этого в митохондрии, разобщая в них дыхание и окислительное фосфорилирование, нарушает синтез белков, поток ионов калия через мембрану клетки и органелл. Это отрицательно влияет на ЦНС.
 Механическая или подпеченочная желтуха развивается вследствие снижения оттока желчи при непроходимости желчного протока – желчные камни, новообразования поджелудочной железы, гельминтозы.
Механическая или подпеченочная желтуха развивается вследствие снижения оттока желчи при непроходимости желчного протока – желчные камни, новообразования поджелудочной железы, гельминтозы.
 В результате застоя желчи происходит растяжение желчных капилляров, увеличивается проницаемость их стенок. Не имеющий оттока в кишечник прямой билирубин поступает в кровь, в результате развивается гипербилирубинемия. Т. к. билирубин в кишечник не попадает, продуктов его катаболизма уроблиногенов в моче и кале нет. Кал обесцвечен.
В результате застоя желчи происходит растяжение желчных капилляров, увеличивается проницаемость их стенок. Не имеющий оттока в кишечник прямой билирубин поступает в кровь, в результате развивается гипербилирубинемия. Т. к. билирубин в кишечник не попадает, продуктов его катаболизма уроблиногенов в моче и кале нет. Кал обесцвечен.
 • В тяжелых случаях, вследствие переполнения гепатоцитов прямым билирубином, реакция конъюгации с глюкуроновой кислотой может нарушаться, присоединяется печеночноклеточная желтуха. В результате в крови увеличивается концентрация непрямого билирубина. • В моче резко увеличен уровень билирубина (цвет темного пива) и снижено количество уробилина, практически отсутствует стеркобилин кала (сероватобелое окрашивание).
• В тяжелых случаях, вследствие переполнения гепатоцитов прямым билирубином, реакция конъюгации с глюкуроновой кислотой может нарушаться, присоединяется печеночноклеточная желтуха. В результате в крови увеличивается концентрация непрямого билирубина. • В моче резко увеличен уровень билирубина (цвет темного пива) и снижено количество уробилина, практически отсутствует стеркобилин кала (сероватобелое окрашивание).
 желтуха – причиной может быть нарушение на всех трех стадиях превращения билирубина") Паренхиматозная (печеночно-клеточная) желтуха – причиной может быть нарушение на всех трех стадиях превращения билирубина в печени: извлечение билирубина из крови печеночными клетками, его конъюгирование и секреция в желчь. Наблюдается при вирусных и других формах гепатитов, циррозе и опухолях печени, жировой дистрофии печени, при отравлении токсическими гепатотропными веществами, при врожденных патологиях.
Паренхиматозная (печеночно-клеточная) желтуха – причиной может быть нарушение на всех трех стадиях превращения билирубина в печени: извлечение билирубина из крови печеночными клетками, его конъюгирование и секреция в желчь. Наблюдается при вирусных и других формах гепатитов, циррозе и опухолях печени, жировой дистрофии печени, при отравлении токсическими гепатотропными веществами, при врожденных патологиях.
 Так как нарушаются все процессы превращения билирубина в печени, гипербилирубинемия развивается за счет обеих фракций, преимущественно прямого билирубина. Концентрация его возрастает из-за нарушения секреции в желчь и увеличения проницаемости мембран клеток печени. Количество непрямого билирубина возрастает за счет функциональной недостаточности гепатоцитов и/или снижения их количества.
Так как нарушаются все процессы превращения билирубина в печени, гипербилирубинемия развивается за счет обеих фракций, преимущественно прямого билирубина. Концентрация его возрастает из-за нарушения секреции в желчь и увеличения проницаемости мембран клеток печени. Количество непрямого билирубина возрастает за счет функциональной недостаточности гепатоцитов и/или снижения их количества.
, умеренно увеличена концентрация уробилина, уровень стеркобилина") В моче определяется билирубин (цвет крепкого черного чая), умеренно увеличена концентрация уробилина, уровень стеркобилина кала в норме или снижен.
В моче определяется билирубин (цвет крепкого черного чая), умеренно увеличена концентрация уробилина, уровень стеркобилина кала в норме или снижен.
 У детей вариантами паренхиматозной желтухи являются физиологическая желтуха новорожденных и недоношенных, желтуха, вызываемая молоком матери. Наследственными формами паренхиматозной желтухи являются синдромы : Ø Жильбера-Мейленграхта, Ø Дубина-Джонсона, Ø Криглера-Найяра.
У детей вариантами паренхиматозной желтухи являются физиологическая желтуха новорожденных и недоношенных, желтуха, вызываемая молоком матери. Наследственными формами паренхиматозной желтухи являются синдромы : Ø Жильбера-Мейленграхта, Ø Дубина-Джонсона, Ø Криглера-Найяра.
 Желтухи часто встречаются у новорожденных Гемолитическая болезнь новорожденного Причины: Несовместимость крови матери и плода по группе или по резус-фактору. Накопление гидрофобной формы билирубина в подкожном жире обуславливает желтушность кожи. Однако реальную опасность представляет накопление билирубина в сером веществе нервной ткани и ядрах ствола с развитием «ядерной желтухи» (билирубиновая энцефалопатия).
Желтухи часто встречаются у новорожденных Гемолитическая болезнь новорожденного Причины: Несовместимость крови матери и плода по группе или по резус-фактору. Накопление гидрофобной формы билирубина в подкожном жире обуславливает желтушность кожи. Однако реальную опасность представляет накопление билирубина в сером веществе нервной ткани и ядрах ствола с развитием «ядерной желтухи» (билирубиновая энцефалопатия).
 Лабораторная диагностика: В крови выявляются выраженная анемия, ретикулоцитоз, эритро- и нормобластоз. Гипербилирубинемия за счет непрямой фракции от 100 до 342 мкмоль/л, в дальнейшем присоединяется и прямая фракция. Уровень билирубина в крови быстро нарастает и к 3 -5 дню жизни достигает максимума.
Лабораторная диагностика: В крови выявляются выраженная анемия, ретикулоцитоз, эритро- и нормобластоз. Гипербилирубинемия за счет непрямой фракции от 100 до 342 мкмоль/л, в дальнейшем присоединяется и прямая фракция. Уровень билирубина в крови быстро нарастает и к 3 -5 дню жизни достигает максимума.
 желтуха новорожденных Причины • относительное снижение активности УДФглюкуронилтрансферазы в первые дни жизни,") Физиологическая (транзиторная) желтуха новорожденных Причины • относительное снижение активности УДФглюкуронилтрансферазы в первые дни жизни, связанное с повышенным распадом фетального гемоглобина, • абсолютное снижение активности УДФглюкуронилтрансферазы в первые дни жизни, • дефицит лигандина, • слабая активность желчевыводящих путей.
Физиологическая (транзиторная) желтуха новорожденных Причины • относительное снижение активности УДФглюкуронилтрансферазы в первые дни жизни, связанное с повышенным распадом фетального гемоглобина, • абсолютное снижение активности УДФглюкуронилтрансферазы в первые дни жизни, • дефицит лигандина, • слабая активность желчевыводящих путей.
 Симптомы исчезают спустя 1 -2 недели после рождения. Лабораторная диагностика: Увеличение концентрации свободного билирубина в сыворотке до 140 -240 мкмоль/л.
Симптомы исчезают спустя 1 -2 недели после рождения. Лабораторная диагностика: Увеличение концентрации свободного билирубина в сыворотке до 140 -240 мкмоль/л.
 Желтуха недоношенных Причины • относительное снижение активности УДФглюкуронилтрансферазы в первые дни жизни, связанное с повышенным распадом фетального гемоглобина, • абсолютное снижение активности УДФглюкуронилтрансферазы в первые дни жизни, • дефицит лигандина, • слабая активность желчевыводящих путей.
Желтуха недоношенных Причины • относительное снижение активности УДФглюкуронилтрансферазы в первые дни жизни, связанное с повышенным распадом фетального гемоглобина, • абсолютное снижение активности УДФглюкуронилтрансферазы в первые дни жизни, • дефицит лигандина, • слабая активность желчевыводящих путей.
 Исчезает спустя 3 -4 недели после рождения. Лабораторная диагностика: Увеличение концентрации свободного билирубина в сыворотке до максимума на 5 -6 дни после рождения, более выражено по сравнению с физиологической желтухой.
Исчезает спустя 3 -4 недели после рождения. Лабораторная диагностика: Увеличение концентрации свободного билирубина в сыворотке до максимума на 5 -6 дни после рождения, более выражено по сравнению с физиологической желтухой.
 Негемолитическая гипербилирубинемия новорожденных, вызываемая молоком матери Встречается у 1% вскармливаемых грудью новорожденных. Причины • Подавление активности УДФглюкуронилтрансферазы, предположительно, эстрогенами материнского молока. Проявляется желтухой, иногда с явлениями поражения ЦНС. Лабораторная диагностика • Увеличение концентрации свободного билирубина в сыворотке.
Негемолитическая гипербилирубинемия новорожденных, вызываемая молоком матери Встречается у 1% вскармливаемых грудью новорожденных. Причины • Подавление активности УДФглюкуронилтрансферазы, предположительно, эстрогенами материнского молока. Проявляется желтухой, иногда с явлениями поражения ЦНС. Лабораторная диагностика • Увеличение концентрации свободного билирубина в сыворотке.
 Синдром Жильбера-Мейленграхта Причины аутосомно-доминантное нарушение элиминации билирубина из плазмы крови, понижение активности УДФглюкуронилтрансферазы. Выявляется в юношеском возрасте и продолжается в течение многих лет, обычно всю жизнь. Наблюдается у 2 -5% населения, мужчины страдают чаще женщин (соотношение 10: 1). Лабораторная диагностика • Периодическое повышение содержание свободного билирубина плазмы, вызванное с провоцирующими факторами.
Синдром Жильбера-Мейленграхта Причины аутосомно-доминантное нарушение элиминации билирубина из плазмы крови, понижение активности УДФглюкуронилтрансферазы. Выявляется в юношеском возрасте и продолжается в течение многих лет, обычно всю жизнь. Наблюдается у 2 -5% населения, мужчины страдают чаще женщин (соотношение 10: 1). Лабораторная диагностика • Периодическое повышение содержание свободного билирубина плазмы, вызванное с провоцирующими факторами.
 Синдром Дубина-Джонсона Причины Аутосомно-доминантная недостаточность выведения конъюгированного билирубина из гепатоцитов в желчные протоки. • Встречается чаще у мужчин, выявляется в молодом возрасте, реже после рождения. Лабораторная диагностика • Увеличение содержания свободного и конъюгированного билирубина в плазме. Характерны билирубинурия, понижение содержания уробилина в кале и моче.
Синдром Дубина-Джонсона Причины Аутосомно-доминантная недостаточность выведения конъюгированного билирубина из гепатоцитов в желчные протоки. • Встречается чаще у мужчин, выявляется в молодом возрасте, реже после рождения. Лабораторная диагностика • Увеличение содержания свободного и конъюгированного билирубина в плазме. Характерны билирубинурия, понижение содержания уробилина в кале и моче.
 Синдром Криглера-Найяра • Тип I Причины Полное отсутствие активности УДФглюкуронилтрансферазы вследствие аутосомно-рецессивного генетического дефекта. Лабораторная диагностика • Гипербилирубинемия появляется в первые дни (часы) после рождения. Характерна интенсивная желтуха. Непрерывное возрастание содержания свободного билирубина в плазме до 200 -800 мкмоль/л (в 15 -50 раз выше нормы). Отсутствие конъюгированного билирубина в желчи.
Синдром Криглера-Найяра • Тип I Причины Полное отсутствие активности УДФглюкуронилтрансферазы вследствие аутосомно-рецессивного генетического дефекта. Лабораторная диагностика • Гипербилирубинемия появляется в первые дни (часы) после рождения. Характерна интенсивная желтуха. Непрерывное возрастание содержания свободного билирубина в плазме до 200 -800 мкмоль/л (в 15 -50 раз выше нормы). Отсутствие конъюгированного билирубина в желчи.
 • Тип II Причины • При синдроме Криглера-Найяра II типа отмечается аутосомно-рецессивный тип наследования. Генетически обусловленное снижение активности УДФглюкуронилтрансферазы. • Желтуха менее интенсивна. Лабораторная диагностика • Содержание непрямого билирубина в крови в 5 -20 раз выше нормы. В желчи есть билирубинглюкуронид.
• Тип II Причины • При синдроме Криглера-Найяра II типа отмечается аутосомно-рецессивный тип наследования. Генетически обусловленное снижение активности УДФглюкуронилтрансферазы. • Желтуха менее интенсивна. Лабораторная диагностика • Содержание непрямого билирубина в крови в 5 -20 раз выше нормы. В желчи есть билирубинглюкуронид.
гемолитическая, обтурационная, печеночно-клеточная. Желтуха новорожденных.
Первая реакция катаболизма гема происходит при участии NADPH-зависимого ферментативного комплекса гемоксигеназы. Ферментная сисгема локализована в мембране ЭР, в области электронтранспортных цепей микросомального окисления. Фермент катализирует расщепление связи между двумя пиррольными кольцами, содержащих винильные остатки, — таким образом, раскрывается структура кольца В ходе реакции образуются линейный тетрапир-рол — биливердин(пигмент жёлтого цвета) и монооксид углерода (СО), который получается из углерода метениловой группы. Гем индуцирует транскрипцию гена гемоксигеназы, абсолютно специфичной по отношению к тему. Ионы железа, освободившиеся при распаде гема, могут быть использованы для синтеза новых молекул гемоглобина или для синтеза других железосодержащих белков. Биливердин восстанавливается до билирубина NADPH-зависимым ферментом биливердинредуктазой. Билирубин образуется не только при распаде гемоглобина, не также при катаболизме других гемсодержащю белков, таких как цитохромы и миоглобин. При распаде 1 г гемоглобина образуется 35 мг билирубина, а в сутки у взрослого человека — примерно 250-350 мг билирубина. Дальнейший метаболизм билирубина происходит в печени.

Билирубин, образованный в клетках РЭС (селезёнки и костного мозга), плохо растворим в воде, по крови транспортируется в комплексе с белком плазмы крови альбумином. Эту форму билирубина называют неконъюгированным билирубином. Каждая молекула альбумина связывает (или даже 3) молекулы билирубина, одна из которых связана с белком более прочно (более высокое сродство), чем другие. При сдвиге рН крови в кислую сторону (повышение концентрации кетоновых тел, лактата) изменяются заряд, конформация альбумина, снижается сродство к билирубину. Поэтому билирубин, связанный с альбумином непрочно, может вытесняться из центров связывания и образовывать комплексы с коллагеном межклеточного матрикса и липидами мембран. Ряд лекарственных соединений конкурирует с билирубином за высокоаффинный, имеющий высокое сродство центр альбумина.
Поглощение билирубина паренхиматозными клетками печени. Комплекс «альбумин-билирубин», доставляемый с током крови в печеНb, на поверхности плазматической мембраны гепатоцита диссоциирует. Высвобожденный билирубин образует временный комплекс с липидами плазматической мембраны. Облегчённая диффузия билирубина в гепатоциты осуществляется двумя типами белков-переносчиков: лигандина (он транспортирует основное количество билирубина) и протеина Z. Активность поглощения билирубина гепатоцитом зависит от скорости его метаболизма в клетке. Лигандин и протеин Z обнаружены также в клетках почек и кишечника, поэтому при недостаточности функции печени они способны компенсировать ослабление процессов детоксикации в этом органе.
Конъюгация билирубина в гладком ЭР. В гладком ЭР гепатоцитов к билирубину присоединяются (реакция конъюгации) полярные группы, главным образом от глюкуроновой кислоты.Билирубин имеет 2 карбоксильные группы, поэтому может соединяться с 2 молекулами глюкуроновой кислоты, образуя хорошо растворимый в воде конъюгат — диглюкуронид билирубина (конъюгированный, или прямой, билирубин).
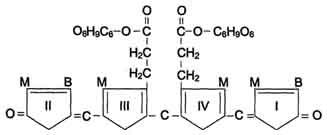
Донором глюкуроновой кислоты служит УДФ-глюкуронат. Специфические ферменты, УДФ-глюкуронилтрансферазы (уридиндифосфоглюкуронилтрансферазы) катализируют образование моно- и диглюкуронидов билирубина.

Секреция конъюгированного билирубина в жёлчь идёт по механизму активного транспорта, т.е. против градиента концентрации. Активный транспорт является, вероятно, ско-рость-лимитирующей стадией всего процесса метаболизма билирубина в печени. В норме диглюкуронид билирубина — главная форма экскреции билирубина в жёлчь, однако не исключается присутствие небольшого количества моноглюкуронида. Транспорт конъюгирован-ного билирубина из печени в жёлчь активируется теми же лекарствами, которые способны индуцировать конъюгацию билирубина. Таким образом, можно сказать, что скорость конъюгации билирубина и активный транспорт билирубинглюкуронида из гепатоцитов в жёлчь строго взаимосвязаны В кишечнике поступившие билирубинглюкурониды гидролизуются специфическими бактериальными ерментами β-глюкуронидазами, которые гидролизуют связь между билирубином и остатком глюкуроновой кислоты. Освободившийся в ходе этой реакции билирубин под действием кишечной микрофлоры восстанавливается с образованием группы бесцветных тет-рапиррольных соединений – уробилиногенов В подвздошной и толстой кишках небольшая часть уробилиногенов снова всасывается, попадает с кровью воротной вены в печеНb. Основная часть уробилиногена из печени в составе жёлчи выводится в кишечник и выделяется с фекалиями из организма, часть уробилиногена из печени поступает в кровь и удаляется с мочой в форме уробилина. В норме большая часть бесцветных уробилиногенов, образующихся в толстой кишке, под действием кишечной микрофлоры окисляется в прямой кишке до пигмента коричневого цвета уробилина и удаляется с фекалиями. Цвет фекалий обусловлен присутствием уробилина.
Причинами гипербилирубинемии могут быть увеличение образования билирубина, превышающее способность печени экскретировать его, или повреждение печени, приводящее к нарушению секреции билирубина в жёлчь в нормальных количествах. Гипербилирубинемию отмечают также при закупорке желчевыводящих протоков печени. Во всех случаях содержание билирубина в крови повышается. При достижении определённой концентрации он диффундирует в ткани, окрашивая их в жёлтый цвет. Пожелтение тканей из-за отложения в них билирубина называют желтухой. Клинически желтуха может не проявляться до тех пор, пока концентрация билирубина в плазме крови не превысит верхний предел нормы более чем в 2,5 раза, т.е. не станет выше 50 мкмоль/л.
Гемолитическая (надпечёночная) желтуха. Известно, что способность печени образовывать глюкурониды и выделять их в жёлчь в 3-4 раза превышает их образование в физиологических условиях. Гемолитическая (надпечёночная) желтуха — результат интенсивного гемолиза эритроцитов. Она обусловлена чрезмерным образованием билирубина, превышающим способность печени к его выведению. Гемолитическая желтуха развивается при исчерпании резервных возможностей печени. Основная причина надпечёночной желтухи — наследственные или приобретённые гемолитические анемии. При гемолитических анемиях, вызванных сепсисом, лучевой болезнью, дефицитом глюкозо-6-фосфатдегидрогеназы эритроцитов, та-лассемией, переливанием несовместимых групп крови, отравлением сульфаниламидами, количество освобождающегося из эритроцитов гемоглобина за сутки может доходить до 45 г (при норме 6,25 г), что значительно увеличивает образование билирубина. Гипербилирубинемия у больных гемолитической желтухой обусловлена значительным повышением (103 — 171 мкмоль/л) в крови концентрации альбуминсвязанного не-конъюгированного билирубина (непрямой билирубин). Образование в печени и поступление в кишечник больших количеств билирубинглюкуронидов (прямой билирубин) ведёт к усиленному образованию и выделению с калом и мочой уробилиногенов и более интенсивной их окраски. Один из главных признаков гемолитической желтухи — повышение содержания в крови не-конъюгированиого (непрямого) билирубина. Это позволяет легко отличить её от механической (подпечёночной) и печёночно-клеточной (печёночной) желтух. Неконъюгированный билирубин токсичен. Гидрофобный, липофильный неконъюгирован-ный билирубин, легко растворяясь в липидах мембраны и проникая вследствие этого в митохондрии, разобщает в них дыхание и окислительное фосфорилирование, нарушает синтез белка, поток ионов калия через мембрану клетки и органелл. Это отрицательно сказывается а состоянии ЦНС, вызывая у больных ряд характерных неврологических симптомов.
Желтуха новорождённых. Частая разновидность гемолитической желтухи новорождённых — «физиологическая желтуха», наблюдающаяся в первые дни жизни ребёнка. Причиной повышения концентрации непрямого билирубина в крови служит ускоренный гемолиз и недостаточность функции белков и ферментов печени, ответственных за поглощение, конъюгацию и секрецию прямого билирубина. У новорождённых не только снижена активность УДФ-глюкуронилтрансферазы, но и, по-видимому, недостаточно активно происходит синтез второго субстрата реакции конъюгации УДФ-глюкуроната. Известно, что УДФ-глюкуронилтрансфераза — индуцируемый фермент. Одно из неприятных осложнений «физиологической желтухи» — билирубиновая энцефалопатия. Когда концентрация неконъюгированного билирубина превышает 340 мкмоль/л, он проходит через гематоэнцефалический барьер головного мозга и вызывает его поражение.
Печёночно-клеточная (печёночная) желтуха. Печёночно-клеточная (печёночная) желтуха обусловлена повреждением гепатоцитов и жёлчных капилляров, например, при острых вирусных инфекциях, хроническом и токсических гепатитах.Причина повышения концентрации билирубина в крови — поражение и некроз части печёночных клеток. Происходит задержка билирубина в печени, чему способствует резкое ослабление метаболических процессов в поражённых гепатоцитах, которые теряют способность нормально выполнять различные биохимические и физиологические процессы, в частности переводить конъюгированный (прямой) билирубин из клеток в жёлчь против градиента концентрации. Для печёночно-клеточной желтухи характерно то, что вместо преобладающих в норме диглюкуронидов билирубина в поражённой печёночной клетке образуются главным образом моноглюкурониды В результате деструкции печёночной паренхимы образующийся прямой билирубин частично попадает в большой круг кровообращения, что ведёт к желтухе. Экскреция жёлчи также нарушена. Билирубина в кишечник попадает меньше, чем в норме. При печёночно-клеточной желтухе повышается концентрация в крови как общего билирубина, так и обеих его фракций — неконъюгированного (непрямого) и конъюгированного (прямого). Так как в кишечник поступает меньше билирубинглюкуронида, то и количество образующегося уробилиногена также снижено. Поэтому кал гипохоличный, т.е. менее окрашенный. Моча, наоборот, имеет более интенсивную окраску за счёт присутствия там не только уробилинов, но и конъюгированного билирубина, который хорошо растворим в воде и экскретируется с мочой.
Механическая, или обтурационная (подпечёночная) желтуха. Механическая, или обтурационная (подпечёночная), желтуха развивается при нарушении желчеотделения в двенадцатиперстную кишку. Это может быть вызвано закупоркой жёлчных протоков, например при желчнокаменной болезни, опухолью поджелудочной железы, жёлчного пузыря, печени, двенадцатиперстной кишки, хроническим воспалением поджелудочной железы или послеоперационным сужением общего жёлчного протока При полной закупорке общего жёлчного протока конъюгированный билирубин в составе желчи не поступает в кишечник, хотя гепатоциты продолжают его вырабатывать. Поскольку билирубин в кишечник не попадает, продуктов его катаболизма уробилиногенов в моче и кале нет. Кал обесцвечен. Так как нормальные пути экскреции билирубина заблокированы, происходит его утечка в кровь, поэтому в крови больных повышена концентрация конъюгированного билирубина. Растворимый билирубин экскретируется с мочой, придавая ей насыщенный оранжево-коричневый цвет.