Проба Амбурже, что показывает анализ мочи, какой метод сбора мочи
Проба Амбурже – это один из видов анализа мочи. Метод Амбурже направлен на выявление количества кровяных тел в биологическом материале. Основная цель анализа заключается в диагностировании почечных заболеваний. Врачам проба Амбурже помогает определить, какая именно почечная болезнь повлекла за собой появление выделенных элементов крови в моче.
На исследование отправляют пациентов с подозрением на хронический пиелонефрит или гломерулонефрит. Оба заболевания являются следствием воспаления почек, и в тяжелой стадии могут перейти на хроническую основу. На раннем этапе развития заболевания мочеиспускательных путей крайне важно получить достоверные результаты исследований. По анализам врачу будет легче назначить лечение.
Проба основывается на количественном анализе форменных элементов в моче. Анализ проводится в центрифуге и камере, подсчитывающей число кровяных тел. Поэтому он крайне точный и многие пациенты своевременно узнают о своих заболеваниях еще на стадии их формирования.
Проба Амбурже
Исследование проводится с целью определения отношения числа лейкоцитов и эритроцитов в моче. Симптомы, свидетельствующие о проблемах в мочеполовой системе, являются основной причиной назначения данного анализа.
Показания к назначению анализа
Следующий ряд симптомов указывает на возможное заболевание почек и мочеполовой системы:
- Температура тела выше нормы.
- Постоянное чувство тошноты, переходящее в рвоту.
- Боли в пояснице.
- Отек конечностей. Для определения степени отека применяется бидигитальная пальпация.
- Утреннее мочеиспускание без большого количества мочи и частые позывы в туалет.
Достоинства и недостатки пробы Амбурже
Преимущества пробы
Основным достоинством пробы является то, что анализ крайне точен в сравнении с другими исследованиями биологического материала. Тест крайне чувствителен и показывает даже слабое, только проявляющее себя заболевание почек.
С результатами анализа можно точно идентифицировать болезнь пациента. Проба идеально подойдет для ранней диагностики заболеваний у детей младшего-дошкольного возраста. Для детей в этот период крайне важно сразу определить заболевание.
Недостатки пробы
Однако, тест совершенно противопоказан онкобольным и лежачим пациентам. Проблема в методе. Получение точных результатов требует от человека ограничить питье, и не опорожняться в течение трех часов. Для тяжелобольных и пациентов, у которых уже наступила запущенная стадия болезни, это почти невыполнимо. Также стоит учесть, что проба – это общее исследование почек.
Если только одна почка подвергнется заболеванию, этот тест не сможет определить, какая именно. Результат покажет менее запущенную степень болезни за счет того, что проверяются одновременно две почки. Из-за невозможности применения данного метода к односторонним заболеваниям мочеполовой системы проба Амбурже становится менее эффективной.
Люди с заболеванием предстательной железы не способны качественно собрать биосубстрат, небольшое количество урины остается в организме. Крайне важен нормальный отток мочи для получения достаточного количества биоматериалов.
Правила сдачи пробы Амбурже
Процесс сдачи анализа мочи по пробе Амбурже многоступенчатый, и подготовка к нему идет в течение трех дней. Это делается для максимальной точности результатов, так как проба крайне чувствительна к микроорганизмам и другими раздражителям в моче.
Подготовка к анализу
Способ сбора порции урины прояснит лечащий врач. Подготовка к анализу мочи по Амбурже происходит следующим образом:
- За сутки до сдачи необходимо ограничить себя в употреблении жидкостей.
- В ночь перед сдачей пробы также запрещается пить.
- Не стоит искусственно создавать позывы в туалет с помощью мочегонных препаратов.
 Лекарство может повлиять на результаты исследования.
Лекарство может повлиять на результаты исследования. - Жидкости с высоким содержанием кофеина и красителей рекомендуется исключить. К этой категории относится кофе, соки, алкоголь.
- Нельзя употреблять острую и кислую пищу в течение 24 часов. Соль также исключена из рациона на период сдачи анализа.
- Во время менструации не стоит сдавать пробу Амбурже, так как это приведет к неточным результатам.
- Перед всеми анализами урины необходимо тщательно мыть половые органы, сдача этого анализа не исключение.
 Лекарство может повлиять на результаты исследования.
Лекарство может повлиять на результаты исследования.Необходимо приобрести стерильную емкость для сбора биоматериала.
Сбор биосубстрата
Для правильного анализа важно соблюсти основные правила сбора урины. Однако, для пробы Амбурже существует специальный порядок действий. Необходимо придерживаться следующих правил во время сбора мочи для пробы:
- Опорожнить мочевой пузырь до конца. Остаток урины может сильно сказаться на результате пробы.
- Записать время опорожнения пузыря.
- Придерживаясь инструкции, необходимо выждать три часа.
- Снова опорожниться, но уже в баночку для сбора анализов. Перед сбором необходимо вымыть наружные половые органы.
- Во время ожидания трех часов перед сбором урины не стоит ограничивать себя в жидкости и пище.

Игнорирование данных правил приведет к ложному анализу и результат не покажет точного ответа на предмет прогрессирования болезней в организме.
Процедура проведения анализа
Пробу производят следующим образом. Баночка с уриной отправляется в лабораторию. Собранный биоматериал помещают в пробирку для центрифугирования в миллилитрах. Это необходимо для того, чтобы осадок и ненужные микроэлементы поднялись, а кровяные клетки осели в пробирке. Вращение центрифуги продолжается пять минут, после чего ее останавливают и извлекают пробирку.На поверхности мочи образуется внешний слой, который снимают. Высота остаточного слоя должна составлять 1 мл. Материал снова помещается в центрифугу и подвергается вращению. Скорость вращения 2 000 оборотов в минуту. Позже, специализированный инструмент осуществляет подсчет кровяных тел в биоматериале.
Скорость вращения 2 000 оборотов в минуту. Позже, специализированный инструмент осуществляет подсчет кровяных тел в биоматериале.
Интерпретация результатов пробы Амбурже
После проведения подсчета больному выдается результат анализа. Итоговые цифры будут показывать количество лейкоцитов и эритроцитов, осевших за одну минуту.
Лаборанты подсчитывают количество клеток в 1 мкл осадка мочи. В случае проведения анализа в камере для подсчета клеток Горячева или Бюркера, количественное соотношение высчитывается по следующей формуле:
x=H/0,9
Н – это количество клеток в камере (отдельно считаются лейкоциты и эритроциты). Число 0,9 отвечает за объем камеры. В зависимости от камеры и ее объема и будет изменяться формула. Но всегда остается неизменным тот факт, что количество клеток делится на общий объем.
Число форменных элементов крови в моче высчитывается по следующей формуле:
A=x*(1000*V)/(S*Т)
А – отвечает за число клеток, которые выделились в моче за минуту вращения в центрифуге.
Х – результат, полученный из предыдущей формулы.
1 000 – объем осадка в кубических миллиметрах.
V – общий объем осадка мочи, которая была выделена за 3 часа.
S – отвечает за то, сколько мл биоматериала находится в центрифуге.
T – время сбора мочи.
Нормальные результаты анализа
Нормой считается показатель 1 000 для эритроцитов и 2 000 для лейкоцитов. Значение может варьироваться. Однако, сильное превышение нормы будет говорить о серьезном заболевании почек. Другой вариант причины плохого результата – это не соблюдены все правила сбора анализов для пробы. При выраженных симптомах и подозрении болезни мочеполовой системы результат необходимо будет закрепить дополнительными исследованиями.
Анализы, соответствующие норме, свидетельствуют о здоровом организме. В том случае, если жалобы на симптомы мочеполовых болезней сохраняются, то проводится второе обследование и назначаются более узконаправленные анализы.
Отклонения от нормы
Повышенное содержание форменных тел говорит о развитии заболевания почек. Значение лейкоцитов и эритроцитов поможет врачам определить, чем именно болен пациент.
Значение лейкоцитов и эритроцитов поможет врачам определить, чем именно болен пациент.
Эритроциты в пробе Амбурже вне нормы
Повышенное содержание эритроцитов в моче и высокий показатель этих кровяных тел, осевших в минуту, говорит о том, что больной страдает от гломерулонефрита. Гломерулонефрит – это острое воспаление почек, который переходит на другие органы при запущенной стадии болезни.
Причинами обычно являются злоупотребление алкоголем, перенапряжение и переохлаждение организма. Заболевания сезонное, и проявляет себя чаще всего зимой и осенью. Именно в это время года люди переохлаждаются. Сырость также может повлиять на здоровье почек.
Возможны и другие причины повышенного содержания эритроцитов. К таким относятся: опухоли, мочекаменная болезнь, поражение организма паразитами, туберкулез и киста.
Лейкоциты в пробе Амбурже вне нормы
Повышенное количество лейкоцитов говорит о таком заболевании, как пиелонефрит. Бактериальное заболевание, которое поражает канальцы почек. Воспаление проявляется болью в пояснице и частыми повышениями температуры тела без видимой на то причины. Основная группа риска заболевания – это женщины от 50 лет. Профилактика данного заболевания заключается в соблюдении личной гигиены и опорожнение мочевого пузыря без длительных задержек.
Воспаление проявляется болью в пояснице и частыми повышениями температуры тела без видимой на то причины. Основная группа риска заболевания – это женщины от 50 лет. Профилактика данного заболевания заключается в соблюдении личной гигиены и опорожнение мочевого пузыря без длительных задержек.
В инфекционные заболевания мочеполовой системы, при которых в урине высокое содержание белых кровяных телец, входят цистит и уретрит. А у мужчин повышенное содержание лейкоцитов – это чаще всего признак простатита.
Если показатель лейкоцитов и эритроцитов понижен, то назначают повторное обследование и снятие пробы Амбурже.
Последствия этих болезней могут стать необратимы для пациента, который вовремя не обратил внимание на здоровье своей мочеполовой системы.
Анализ мочи по амбурже: как собрать, преимущества
Исследованием, помогающее врачам определить количество форменных частей крови в моче для диагностирования почечных заболеваний, является проба Амбурже.
Подготовка к анализу мочи по Амбурже
Для правильного проведения анализа мочи по методу Амбурже, пациенту следует ограничить объем питья в день перед сдачей, а также совсем не употреблять жидкость ночью. Кроме того, нужно придерживаться перечня ограничений, как при общем анализе:
- Не пить мочегонные напитки и продукты, которые способны повлиять на цвет мочи.
- За полдня до сбора анализов нельзя употреблять соленую и пряную пищу, алкоголь.
- Не рекомендуется сдавать мочу женщинам в период менструации.
Вернуться к оглавлениюСбор мочи для любого вида анализа проводится утром. Стоит заранее приобрести в аптеке одноразовый контейнер для мочи, так как чистая и стерильная посуда без следов посторонних веществ очень важна для корректных результатов исследования.
Как собрать материал?
При исследовании этим методом больной ограничивает прием жидкости днем и исключает ночью.Для сбора материала на пробу Амбурже стоит придерживаться общих правил сдачи мочи для исследования. Необходимо соблюдать личную гигиену — непосредственно перед сбором урины вымыть наружные половые органы. При этом нельзя пользоваться моющими средствами. Для сбора материала по методу Амбурже, пациенту после воздержания от питья, необходимо полностью опорожнить мочевой пузырь. Затем нужно отметить время, подождать ровно 3 часа, не справляя малую нужду, и только тогда собрать мочу на анализ. На протяжении этого времени нет никаких ограничений в употреблении пищи или напитков. Рекомендуется сдавать для исследования среднюю порцию струи. После сбора материала, необходимо незамедлительно доставить пробу в лабораторию. Если необходимо взять мочу у маленького ребенка, применяют специальный мочесборник.
Рекомендуется сдавать для исследования среднюю порцию струи. После сбора материала, необходимо незамедлительно доставить пробу в лабораторию. Если необходимо взять мочу у маленького ребенка, применяют специальный мочесборник.
Нормальные значения
Исследование мочи по методу Амбурже представляет собой подсчет количества лейкоцитов и эритроцитов, осевших за одну минуту, и сопоставление результатов с нормой. Нормальными показателем красных кровяных телец считается 1000. Если этот показатель повышен, это свидетельствует о наличии хронического гломерулонефрита или недуга, сопровождающегося гематурией (присутствием крови в моче). Число лейкоцитов, выделяемых в минуту в моче здорового человека равно 2000. Если этот показатель превышен, то пациент страдает хроническим воспалением мочеиспускательных путей и пиелонефритом. В некоторых источниках указана норма лейкоцитов 2500, а эритроцитов — 2000.
Вернуться к оглавлениюОтклонения от нормы
Эритроциты
Повышенное количество эритроцитов в моче свидетельствует о воспалительном заболевании.
Если при анализе Амбурже было выявлено повышенное выделение эритроцитов, это свидетельствует о гломерулонефрите. Это острое воспалительное заболевание, сначала поражающее клубочковый аппарат почек, а затем и остальные системы. Эту болезнь чаще всего вызывает стрептококк. Развитие хронического гломерулонефрита способствует чрезмерное употребление алкоголя, воздействие низких температур и интенсивные физические нагрузки. Кроме того, этот недуг может появиться как следствие перенесенных операций. Чаще всего от гломерулонефрита страдают мужчины. Заболевание носит сезонный характер и проявляется в период холода и сырости. Пациент может страдать от других заболеваний мочеиспускательных путей, которые сопровождаются патологическим выделением эритроцитов.
Вернуться к оглавлениюЛейкоциты
Когда проба Амбурже показала, что количество выделяемых лейкоцитов превышает 2000 за 60 секунд, то пациент страдает хроническим воспалением мочеиспускательных путей и пиелонефритом. Это воспалительный процесс, затрагивающий ткань почек. Женщины в возрастной категории после 50-ти лет наиболее подвержены этому недугу, а также мужчины, которые перенесли травму позвоночника или воспаления мочеполовых систем. Хронический пиелонефрит — частое последствие болезней, при которых нарушен нормальный отток мочи. При пониженном показателе лейкоцитов назначаются повторные анализы и дополнительные провокационные тесты.
Женщины в возрастной категории после 50-ти лет наиболее подвержены этому недугу, а также мужчины, которые перенесли травму позвоночника или воспаления мочеполовых систем. Хронический пиелонефрит — частое последствие болезней, при которых нарушен нормальный отток мочи. При пониженном показателе лейкоцитов назначаются повторные анализы и дополнительные провокационные тесты.
Процедура анализа
Аппарат предназначен для подсчета количества эритроцитов и лейкоцитов.Для подсчета форменных кровяных компонентов в моче, лаборант отправляет материал в центрифугу при 2000 оборотов на минуту. После этого снимается верхний слой, а осадок с небольшим количеством жидкости над ним подлежит исследованию в специальной счетной камере. Там отдельно подсчитывают количество эритроцитов и лейкоцитов. Если лаборант использует камеру Горяева или Брюкера, то расчет происходит по формуле: Х=Н/0,9, где Х — количество клеток в 1 мкл мочи, Н — значение насчитанных в камере клеток, а 0,9 размер самой камеры. Если подсчет кровяных элементов проводился в камере Фукс-Розенталя, то значение 0,9 в формуле следует заменить на 3,2, согласно размерам этой камеры.
Если подсчет кровяных элементов проводился в камере Фукс-Розенталя, то значение 0,9 в формуле следует заменить на 3,2, согласно размерам этой камеры.
Вернуться к оглавлениюДля подсчета количеств лейкоцитов и эритроцитов могут использоваться такие виды камер: Фукс-Розенталя, Горяева или Брюкера.
Преимущества и недостатки
Главным преимуществом анализа мочи по Амбурже считается его точность в сравнении с другими способами подсчета форменных элементов крови. Этот способ позволяет выявить скрытые и вялотекущие заболевания почек и мочеиспускательных путей. С помощью этого анализа, врач может следить за динамикой развития болезни и должным образом корректировать лечение. Метод Амбурже удобно применять для взятия пробы у маленьких детей.
К недостаткам метода Амбурже можно отнести ограничения в употреблении жидкости и мочеиспускании, что может вызывать дискомфорт у пациента. Некоторые заболевания сопровождаются резкими болями и человеку трудно сдерживать позывы к малой нужде. Больные, страдающие от рака предстательной железы и аденомы неспособны до конца опустошать мочевой пузырь. Исследование по методу Амбурже бесполезно при одностороннем поражении почек, так как этот анализ покажет общий результат, который будет занижен.
Больные, страдающие от рака предстательной железы и аденомы неспособны до конца опустошать мочевой пузырь. Исследование по методу Амбурже бесполезно при одностороннем поражении почек, так как этот анализ покажет общий результат, который будет занижен.
Анализ мочи по Амбурже — что это, кому назначается, отклонения
Моча сама по себе информативный материал. Она позволяет сразу же выявить у человека начало заболевания и развитие более серьёзных патологий. Для диагностирования болезней используются различные методы исследования биоматериала. Одним из таких информативных является метод Амбурже. Количественные пробы позволяют выявить лейкоциты, эритроциты и цилиндры. Как правильно собрать материал и норма значений, узнаем в статье.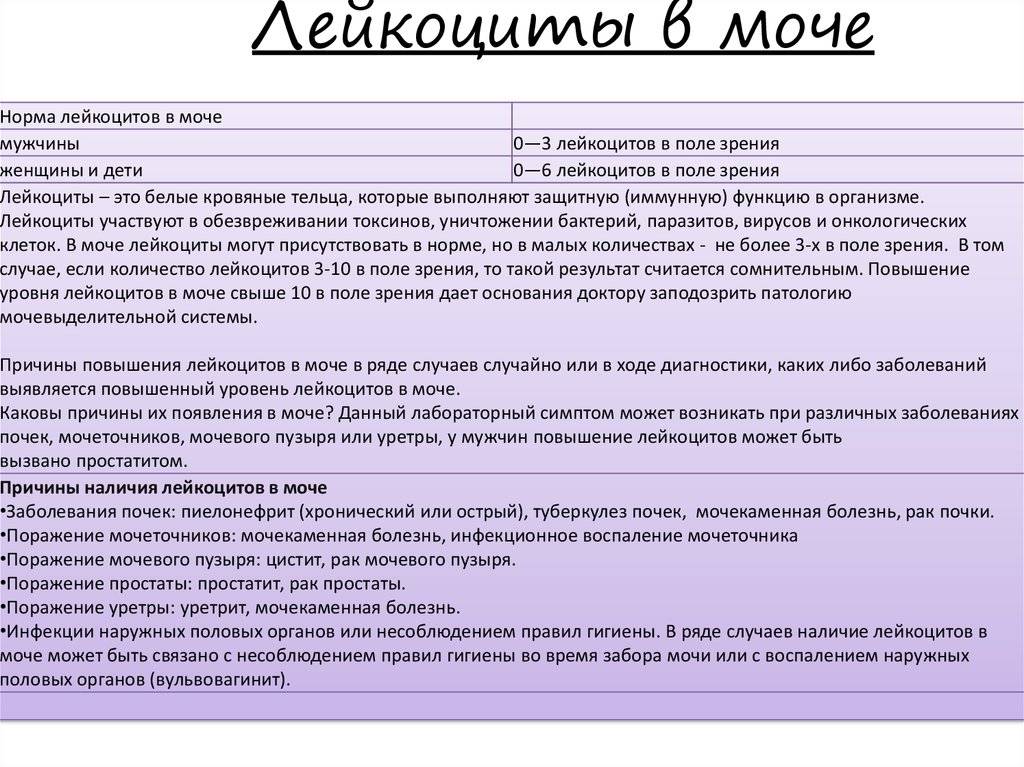
Содержание статьи
Проба Амбурже — назначение
Используя метод Амбурже, врачи могут по количественным значениям крови в урине диагностировать состояние почек. Анализ позволяет очень точно отличить пиелонефрит от гломерулонефрита. Соотношение лейкоцитов и эритроцитов методом Амбурже проводится за одну минуту. Назначают данный метод, людям которые обращаются к врачу с такими симптомами:
- Повышение температуры тела.
- Озноб, рвота.
- Боль в боку или поясничной области.
- Отечность конечностей.
- Частые позывы к мочеиспусканию.
Эти симптомы явно свидетельствую о проблемах в мочеполовой сфере, и чтобы поставить более точный диагноз и выявить причину заболевания, используют метод Амбурже. Но для достоверного и информативного результата, нужно правильно подготовится в забору материала.
А также в анализе не допускаются ошибки при подсчете микроорганизмов, так как это сказывается на диагнозе очень негативно. Несмотря на то, что этот метод очень точный, он имеет ряд недостатков, которые стоит учитывать.
Правила подготовки к забору материала
Для исследования анализа мочи методом Амбурже, больному рекомендуют придерживаться строгих правил:
- За день до проведения анализа, стоит ограничить прием жидкости.
- Ночью перед анализом нельзя пить.
- Не принимать мочегонные препараты.
- Соки, чай, кофе стоит исключить из рациона, за три дня до анализа.
- За сутки не стоит употреблять алкоголь.
- Также за 24 часа до забора материала, исключить из питания соленую, острую и кислую пищу.
- Женщинам в период менструации не рекомендуется проводить анализ по Амбурже.
- Непосредственно перед самым сбором урины, стоит промыть половые органы.
В аптеке необходимо заранее приобрести емкость, куда будет собираться материал. Не стоит использовать домашнюю посуду, так как там могут остаться частички веществ, которые способны помешать во время исследования.
После того, как предварительно пациент подготовился, необходимо утром опорожнить мочевой пузырь полностью. И выполнять ряд действий:
- Отметить время после опорожнения.
- Строго выждать три часа и после этого собирать урину.
- На протяжении трех часов перед сбором материла, не ограничивать себя в еде и питье.
Как только время ожидания приблизиться к трёхчасовой отметке, стоит провести процедуру личной гигиены и приступить непосредственно к забору материала. Первую струю урины стоит слить, а вот для исследования брать только средний поток. Как только материал будет собран его сразу же стоит отнести в лабораторию.
Расшифровка анализа — норма
Тест Амбурже позволяет произвести количественный подсчёт клеток лейкоцитов, эритроцитов и сопоставить их с нормой.
Мочу хорошо перемешивают в специализированной центрофужной пробирке. Центрифуга крутиться около пяти минут, после чего осторожно снимают внешний слой материала, и оставляют только 1 миллилитр с осадком. Затем этот материал снова перемешивается и помещается в камеру для подсчёта клеток лейкоцитов и эритроцитов.
В зависимости от камеры, могут отличаться и единицы измерения – камера Горяева считает в МКЛ, а камера Фукс-Розенталя в куб. мм.
Норма показателей в таблице:
| Клетки | Норма |
|---|---|
| Лейкоциты | 2 000 – 2 500 |
| Эритроциты | 1 000 – 2 000 |
Показания могут быть разными в зависимости от камеры, с помощью которой производился подсчёт клеток. Как правило, на бланке лаборатории указывается нормальные значения и ед. измерения в этой лаборатории.
Отклонения от нормы — чем это грозит больному
Конечно, любой проводимый анализ, позволяет определить заболевание, которое привело пациента в больницу. Отклонения от нормы тех или иных показателей, позволяет докторам поставить диагноз и назначить лечение. Если результаты анализа существенно отличаются от нормы, то пациенту назначаются дополнительные анализы, более узкоспециализированные.
О чем же говорят отклонения от нормы по методу Амбурже? Если было выявлено повышенное количество эритроцитов, это может свидетельствовать о заболевании гломерулонефром. Оно представляет собой острый воспалительный процесс, который поражает клубочки почек, а затем перекидывается на другие системы.
Развитию данного заболевания способствует злоупотребление алкогольных напитков, тяжелые физические нагрузки, переохлаждение. Кроме этого гломерулонефром может возникнуть после перенесенной операции.
Справочно. Этим патологиям больше подвержены представители сильной половины человечества. Как правило, это заболевание носит сезонный характер, и часто поражает людей в холодное и сырое время года.
Повышение лейкоцитов, характеризуется воспалением мочеполовой сферы и хроническим пиелонефритом. Воспаление затрагивает почки, что вызывает боль в пояснице. К группе риска относятся женщины старше пятидесяти лет, а также сильная половина Земли, не так давно перенесших позвоночную травму.
Преимущества и недостатки метода Амбурже
Основным преимуществом метода, является его стопроцентная точность по сравнению с другими исследованиями. Тест показывает все скрытые и слабо текущие заболевания почек и мочеполовой сферы. Используя метод Амбурже, врач может следить за развитием заболевания, и своевременно корректировать лечение.
Часто такой метод применяют для исследования биоматериала у детей. Так как точность исследования позволяет выявить заболевание на ранних стадиях.
Но к сожалению, есть и недостатки, поэтому такое исследование не назначают лежачим и онкобольным людям.
Не каждый больной человек сможет ограничивать себя в питье, и затем выдержать три часа без мочеиспускания. Для больных, стадия заболевания которых находится в острой или хронической форме, не справлять нужду три часа, невыполнимое условие. Онкобольные предстательной железы или аденомы, не способны полностью испражняться.
Методика Амбурже абсолютно неэффективна при одностороннем заболевании почек, так как показывает общее состояние, и соответственно значения будут значительно снижены.
Каковского-Аддиса проба — это… Что такое Каковского-Аддиса проба?
- Каковского-Аддиса проба
— количественный метод исследования мочи, заключающийся в сборе мочи, выделенной за 10-12 часов, определении числа эритроцитов, лейкоцитов и цилиндров в осадке.
Источник: «Медицинская Популярная Энциклопедия»
Медицинские термины. 2000.
- каверна
- кальцитонин
Смотреть что такое «Каковского-Аддиса проба» в других словарях:
Каковского-Аддиса проба — (А. Ф. Каковский, отеч. врач; Th. Addis, 1881 1949, амер. врач) количественный метод исследования мочи, заключающийся в сборе мочи, выделенной за 10 12 часов, определении с помощью счетной камеры числа эритроцитов, лейкоцитов и цилиндров в осадке … Большой медицинский словарь
Проба Каковского — Проба Каковского Аддиса количественный метод исследования мочи, заключающийся в сборе мочи, выделенной за 10 12 часов, определении с помощью счётной камеры числа эритроцитов, лейкоцитов и мочевых цилиндров в осадке небольшой пробы и… … Википедия
Аддиса-Каковского проба — (Th. Addis, 1881 1949, амер. врач; А. Ф. Каковский, отеч. врач) см. Каковского Аддиса проба … Большой медицинский словарь
А́ддиса — Како́вского про́ба — (Th. Addis, 1881 1949, американский врач; А.Ф. Каковский, отеч. врач) см. Каковского Аддиса проба … Медицинская энциклопедия
Аддис, Томас — Томас Чалмерс Аддис (англ. Thomas Chalmers Addis; 1881 1949) американский врач, учёный, педагог, политический деятель, именем которого названа проба по исследованию мочи (проба Каковского Аддиса). Содержание 1 Биография 2 Научная д … Википедия
Обследование больного — I Обследование больного Обследование больного комплекс исследований, направленных на выявление индивидуальных особенностей больного, установление диагноза болезни, обоснование рационального лечения, определение прогноза. Объем исследований при О … Медицинская энциклопедия
Лейкоцитурия — I Лейкоцитурия (leucocyturia; лейкоцит[ы] + греч. uron моча) повышенное по сравнению с нормой выделение лейкоцитов с мочой, признак воспалительного процесса в мочевой системе. Л. несомненна, если при микроскопии осадка мочи обнаруживают более 20… … Медицинская энциклопедия
По́чки — (renes) парный экскреторный и инкреторный орган, выполняющий посредством функции мочеобразования регуляцию химического гомеостаза организма. АНАТОМО ФИЗИОЛОГИЧЕСКИЙ ОЧЕРК Почки расположены в забрюшинном пространстве (Забрюшинное пространство) на… … Медицинская энциклопедия
Коберт, Рудольф — Рудольф Эдвард Коберт Rudolf Eduard Kobert Рудольф Эдвард Коберт (нем … Википедия
Памятка по сбору анализов мочи
Техника сбора мочи на общий анализ
После туалета наружных половых органов для общего анализа собирают утреннюю мочу в одноразовый аптечный контейнер не менее 50 мл.
Контейнер с мочой необходимо доставить в лабораторию с 7-30 до 10 часов утра.
Техника сбора мочи анализа «по Нечипоренко»
После туалета наружных половых органов собирают СРЕДНЮЮ порцию утренней мочи объемом не менее 20 мл. в одноразовый аптечный контейнер первая и последняя порция мочи сливаются в унитаз.
Контейнер с мочой необходимо доставить в лабораторию с 7-30 до 10 часов утра.
Техника сбора суточной мочи
Моча собирается в течение 24 часов. Утром перед сбором мочи провести туалет наружных половых органов. Первая утренняя порция мочи выпускается в унитаз и отмечается время мочеиспускания. Далее, вся моча в течение суток (до того же времени следующего дня) должна собираться в одну ёмкость, (обычно используется 3-х литровая банка с крышкой, которая хранится в холодильнике).
По окончании сбора материала необходимо самостоятельно оценить общий объём собранной мочи. После чего моча перемешивается и часть её (не менее 100 мл) отливается в специальный контейнер. Именно этот контейнер доставляется в лабораторию (весь объём мочи доставлять не надо) с 7-30 до 10 часов утра.
При сдаче мочи в лабораторию необходимо указать с какого по какое время собиралась моча; каков получился общий объем мочи.
Допускается собирать мочу за 12 час.
Техника сбора мочи на бактериологический посев
Подготовить стерильный контейнер для сбора мочи. Вымыть руки с мылом, затем тщательно вымыть наружные половые органы.
после подмывания взять стерильный контейнер для сбора мочи, открыть крышку так, чтобы не касаться внутренней поверхности крышки и контейнера;
выделить первую струю мочи в унитаз, задержать мочеиспускание, подставить контейнер, выделить мочу в контейнер в количестве не менее 10 – 15 мл, оставшуюся порцию мочи спустить в унитаз. Аккуратно закрыть крышкой контейнер, не касаясь краев горлышка.
Не допускается переохлаждение, перегревание и пролив мочи!
Принимается анализ строго с 7:30 до 8:45 (кроме выходных дней)
Техника сбора мочи на глюкозурический профиль
Собирается суточная моча в три чистые отдельные банки, на каждой банке подписывается время сбора мочи.
с 6-00 до 14-00.
с 14-00 до 22-00.
с 22-00 до 6-00.
Измеряется количество собранной мочи в мл.
Измеряется количество собранной мочи.Указывается количество в мл.
Отливается моча из каждой банки в отдельный пластиковый контейнер (не менее 50 мл.), на каждом контейнере подписывается время сбора мочи.
Контейнеры с мочой необходимо доставить в лабораторию с 7-30 до 10 часов утра (кроме выходных дней).
Техника сбора мочи для пробы Реберга
Проба Реберга предполагает забор образцов мочи и крови. Накануне исключить физические нагрузки, спиртные напитки, крепкий чай, острую и жирную пищу.
По окончании сбора материала надо самостоятельно оценить общий объём собранной мочи. После чего моча перемешивается и часть её (не менее 150 мл) отливается в специальный контейнер. Именно этот контейнер доставляется в лабораторию (весь объём мочи доставлять не надо) с 7-30 до 10 часов утра.
При сдаче мочи в лабораторию необходимо указать с какого по какое время собиралась моча и общий объем мочи. Указывается свой рост, вес и количество жидкости, выпитой в течении суток.
При доставке анализа мочи в лабораторию необходимо в этот день сдать кровь для определения уровня креатинина (натощак)
Техника сбора мочи по Зимницкому
В ходе исследования определяют суточный, дневной и ночной диурез и относительную плотность мочи в каждой из 3-х часовой порции
Сбор мочи для пробы Зимницкого производится в определенные часы в течение суток. Проба проводится при обычном питьевом режиме и питании.
Для того, чтобы правильно собрать требуемый материал необходимы:
8 чистых емкостей с крышкой объемом 200 мл каждая – их можно получить в лаборатории больницы ( на каждой подписать время сбора мочи )
Часы, желательно с будильником (сбор мочи должен происходить в определенные часы)
Блокнот для записи потребляемой в течение суток жидкости (в том числе и объем жидкости поступающей с супом, молоком и т.д. )
Как собирать мочу для исследования по Зимницкому?
В 6 часов утра необходимо опорожнить мочевой пузырь в унитаз. В течение всего дня каждые 3 часа необходимо опорожнять мочевой пузырь в баночки (если одной емкости не достаточно, берется дополнительная емкость)
Строго соблюдать 3-часовой разрыв между порциями мочи!
- с 6 до 9 утра — всю мочу, выделенную в течение последующих 3-х часов собрать в первую емкость;
- с 09 до 12 часов – во вторую емкость;
- с 12 до 15 часов — в третью;
- с 15 до 18 часов – в четвертую;
- 18 до 21 часов — в пятую;
- с 21 до 24 часов – в шестую;
- с 24 до 03 часов – в седьмую;
- с 03 до 06 часов утра заполнить последнюю, восьмую емкость.
Наполняемые баночки необходимо содержать в холоде в закрытом виде (в холодильнике).
Утром следующего дня необходимо отнести все баночки с содержимым в лабораторию, дополнительно отдав записи о потребленной в течение суток жидкости.
Общеклинический анализ мочи — сдать в Едином медицинском центре в СПб
Общеклиническое исследование мочи – одна из самых важных диагностических процедур. Ее цель – определение химических и физических характеристик продукта, выделяющегося в результате человеческой жизнедеятельности – мочи.
Микробиологический и химический состав мочи – обязательная составная часть диагностики. Любые отклонения от нормы сговорят о нарушении процессов обмена в организме.
Своевременное исследование мочи помогает диагностировать:
- Нефриты и опухолевые процессы в почках;
- Заболевания предстательной железы;
- Мочекаменную болезнь;
- Патологии мочевого пузыря.
Общий анализ мочи – процедура обязательная, и назначают ее всем пациентам, которые приходят на прием к врачу, при любом диагнозе.
Полученные результаты исследования позволяют делать точные заключения. Общий анализ мочи, наряду с анализами кала и крови используются врачами для подтверждения или опровержения предположений о наличии самых разных болезней.
Собранный биологический материал исследуют в лаборатории в полном соответствии с утвержденными нормами.
Анализ мочи по Нечипоренко
Исследование специальное, используется для определения состояния и функциональности почек и мочевыводящих путей. Обычно этот анализ назначают после получения результатов общего анализа, если его результаты не соответствуют нормам. Исследование по Нечипоренко дает более полную картину нарушений и помогает правильно диагностировать состояние пациента.
Проба Реберга-Тареева
Это исследование дает возможность оценить всасывающую и выделительную способность почечных канальцев и диагностировать уровень выделительной способности почек. Для проведения данного исследования необходимо: суточная моча, венозная кровь в день сдачи анализа, данные о возрасте, росте и весе пациента. Пациенту проводится предварительный инструктаж по сбору суточной мочи.
Анализ мочи на сахар
Проверка уровня глюкозы в моче – одна из разновидностей анализов. В норме в моче не должно быть сахара. Если же анализ покажет его присутствие, врач может сделать вывод и о наличии серьезных проблем со здоровьем: это может быть заболевание почек или диабет.
Цены на исследование мочи
Единый диагностический центр проводит клиническое исследование мочи. Наличие собственной лаборатории, оснащённой современной аппаратурой, позволяет получить точный результат в самые короткие сроки.
Цены исследования — доступны. Сделать анализ можно без очереди, и в самые короткие сроки получить точный результат.
Правила сбора общего анализа мочи:
- После пробуждения пациент проводит туалет наружных половых органов. Собирается вся утренняя порция мочи, перемешивается, отливается не менее 50 мл мочи в контейнер и доставить в лабораторию.
Правила сбора пробы Нечипоренко:
- После пробуждения пациент проводит туалет наружных половых органов. Собирается утренняя средняя порция мочи, не менее 50 мл мочи в контейнер и доставить в лабораторию. Обратите внимание!!!! Нельзя собирать мочу во время менструации и в течение 5-7 дней после цистоскопии.
Правила сбора пробы Реберга:
- После пробуждения, при первом утреннем мочеиспускании полностью опорожнить мочевой пузырь. Далее собирать мочу в контейнер со второго мочеиспускания, время мочеиспускания необходимо обязательно записать. Собрать всю мочу, полученную в течении дня и ночи. Утреннюю порцию мочи следующего дня (до времени отмеченного накануне) собрать в контейнер. На протяжении всего периода сбора мочи контейнер хранить в холодильнике при температуре +2+4 С. Собранную мочу тщательно перемешивают, отливают не менее 50 мл, маркируют, указывают суточный диурез, количество выпитой жидкости, рост , вес пациента , доставляют в лабораторию.
Исследуя мочу, специалисты определяют основные характеристики, в числе которых следующие параметры:
- кислотность;
- цвет;
- пенистость и прозрачность;
- запах;
- удельный вес материала;
- количество белков, глюкозы, кровяные тельца;
- цилиндры, бактерии, грибы;
- моли.
Если возникает необходимость хранить мочу некоторое время, следует выбрать темное прохладное место.
Требуется предоставить полученный материал на исследование в течение полутора часов. Данный анализ – обычная процедура при плановых осмотрах. Мы предлагаем пройти исследование частным образом.
Диагноз не может быть поставлен на основании только анализа мочи, так как одинаковые показатели могут соответствовать различным болезням.
Метод Амбурже. Всё, что нужно знать о своих анализах. Самостоятельная диагностика и контроль за состоянием здоровья
Читайте также
Мой метод
Мой метод Я твёрдо решил бросить курить и рассчитывал добиться результата сам, без сложных подготовок и специальных курсов. У меня это получилось. И поверьте — всё оказалось значительно проще, чем я думал.После того, как я перепробовал разные методы бросить курить, я
Нехирургический метод
Нехирургический метод Сколько Ольга себя помнила, у нее всегда был насморк. Но в молодости на это не обращаешь особого внимания. А вот с годами насморк стал докучать женщине. Тем более что он стал сопровождаться сильной головной болью и заложенностью в носу. Никакие
Оперативный метод
Оперативный метод Еще совсем недавно самым распространенным способом извлечения камня (если он не отходит, конечно, сам) считался оперативный метод. Больной перед операцией должен пройти полное клиническое обследование. Операция обычно длится 2–3 часа. Период
Метод И. Добротворского
Метод И. Добротворского Очень хорошо на больных хроническим пиелонефритом действует схема лечения, предложенная врачом Игорем Доб-ротворским: очищение почек, фитотерапия, исцеляющие настои, дыхательная гимнастика, медленный бег, обливание прохладной водой, хождение
Метод Стрельниковой
Метод Стрельниковой Данная гимнастика основана на динамичных дыхательных упражнениях, которые сопровождаются движениями рук, ног и туловища. Упражнения дыхательной гимнастики следует выполнять точно и регулярно. Лишь в этом случае комплекс упражнений окажет
Метод Бутейко
Метод Бутейко © ООО «Издательство АСТ»Все права защищены. Никакая часть электронной версии этой книги не может быть воспроизведена в какой бы то ни было форме и какими бы то ни было средствами, включая размещение в сети Интернет и в корпоративных сетях, для частного и
«Полюбившийся метод»
«Полюбившийся метод» Марию Григорьевну больные прозвали «Королевой». Она хорошо известна в клинике. Появление её всегда встречают возгласами: «А, Мария Григорьевна! Опять к нам! Ну, как поживаете?» Она со всеми здоровается, всех знает по имени и отчеству, сестер зовет
МЕТОД ЛЕЧЕНИЯ
МЕТОД ЛЕЧЕНИЯ Практика Доктора Коновалова основана на его способности управлять тонкими энергиями, создавая в лечебном зале своеобразный купол, заполняя все внутреннее пространство зала этой энергией, на много порядков превышающей обычный биоэнергетический фон
ДАОСИСТСКИЙ МЕТОД
ДАОСИСТСКИЙ МЕТОД Основанная на даосистской метафизике, эта восточная философия являет собой сочетание духовной концентрации и манипуляцию половыми органами. В соответствии с требованиями даосистской сексуальности нужно добиться ощущения «ладони в перчатке», чтобы
МЕТОД «ДОЕНИЯ»
МЕТОД «ДОЕНИЯ» В августовском номере журнала «Плейбой» за 1989 год была опубликована статья Донны Пауэлл «Как сделать его больше», в которой она давала рекомендации, как увеличить размеры мужского полового органа. Это очень простой метод, основанный на принципе «доения»,
МЕТОД SE-101
МЕТОД SE-101 Этот метод, известный также под более пространным название: «16 недель естественного развития пениса по плану доктора Тейлора», включает воображение пациента, массаж и надавливание определенных точек (метод шиацу). В рекламной брошюре сообщается, что этот метод
МЕТОД «V»
МЕТОД «V» Несколько лет назад у жителя штата Нью-Джерси Юджина Вискионе возникла не поддающаяся лечению болезнь пейрония (аномальное искривление пениса). Такой синдром может возникнуть в любое время и обычно связывается с травмой члена, особенно при половом
Метод №4: Шок
Метод №4: Шок Этот метод основывается на разрушении ее привычных стереотипов.В этом случае вы говорите с железными интонациями Арнольда Шварценеггера в роли Терминатора:«Непривлекателен? А разве кто-то говорил, что я должен быть привлекателен для тебя? Меня это мало
Метод релаксации 2
Метод релаксации 2 Лягте на пол, руки вдоль туловища, ноги слегка раздвинуты.Сконцентрируйте свое внимание на пальцах ног, направьте «луч» внимания именно на это место, скажите себе: «Мое внимание на пальцах ног, я спокойна, пальцы ног расслаблены»; затем двигайтесь снизу
МЕТОД М. В. КУТУШОВА
МЕТОД М. В. КУТУШОВА Автор собственной теории возникновения рака предлагает и свои методы диагностики и лечения злокачественных новообразований. Будучи опытным врачом-практиком и исследователем, автор прибегает к помощи буддийской философии и тибетской медицины. Его
Анализ мочи по методу Амбурже Правила сбора, норма и результаты
Исследование мочи по методике Т. Амбурже — один из методов определения количества эритроцитов и лейкоцитов в минутном объеме мочи.
Методика была предложена Т. Амбурже в 1954 г. как модификация пробы Каковского-Аддиса и в настоящее время используется в практической медицине при диагностике патологии почек и мочевыделительной системы в целом.
Необходимость таких методов продиктована тем, что при обычном исследовании осадка мочи не учитываются его количество, объем, а также количество взятой на центрифугирование мочи и охватываемое ею поле зрения. микроскоп.
Этот метод учитывает вышеперечисленные показатели и, как следствие, исключает возможные ошибки.
Правила сбора
Неотъемлемой частью любого медицинского исследования является этап подготовки, от которого зависит качество получаемых результатов (поэтому при отсутствии туалета половых органов возможны искажения истинного количества лейкоцитов) .
Назначение данного анализа определяет лечащий врач. Он также уполномочен объяснять суть проводимого исследования и давать пациенту инструкции о том, как подготовиться к сбору мочи.За день до исследования пациенту необходимо ограничить прием жидкости, потребляемой днем, и полностью исключить в ночное время.
Утром необходимо провести гигиеническую обработку наружных половых органов и опорожнить мочевой пузырь (это делается физиологическим способом или введением в него катетера), отметить время и собрать мочу для исследования в чистую банку на 3 часа.
Методика проведения
Исследование проводится в клинической лаборатории врачом-лаборантом.
Его реализация включает в себя несколько этапов:
- Удаление 3 мл мочи, собранной за 10-часовой период.
- Поместите в центрифугу на 5 минут при 2000 об / мин.
- Возьмите надосадочную жидкость так, чтобы на дне остался 1 мл осадка.
- Перемешайте осадок и заполните им счетную камеру, считая лейкоциты и эритроциты в 1 мкл.
Клетки рассчитываются по формуле: Q = (x 1000 v) / st, где:
- Q — количество клеток в объеме полученной мочи за минуту,
- x — количество подсчитанных клеток в камере (отдельно для лейкоцитов и эритроцитов)
- с — количество взятой на центрифугирование мочи (10)
- v — объем мочи, полученной за 3 часа.
- t — время, в течение которого была собрана моча (3 часа = 180 минут).
Полученное число в 1 мкл, умноженное на 1000 (1 мл = 1000 мкл).
Нормы показателей
Независимо от того, какое исследование проводится, для интерпретации его результатов необходимо знать показатели, характерные для здорового человека в тех же условиях, поскольку без этих знаний диагностическое значение полученные результаты теряются.
И поэтому ученые определили, что у человека со здоровыми почками и мочевыводящей системой в анализе мочи по Амбуржу обнаруживается осадок:
- 2000 лейкоцитов.
- 1000 эритроцитов.
Допустимые колебания цифр возможны в пределах 10%.
Оценка результатов
Результаты теста (количество лейкоцитов и эритроцитов) фиксируются по направлению, поступившему в лабораторию вместе с мочой, и отправляются лечащему врачу. Он на основании лабораторных данных и знания нормальных показателей оценивает полученные результаты, сравнивая норму и информацию из лаборатории.
Это помогает врачу диагностировать такие заболевания, как:
- Пиелонефрит — инфекционно-воспалительная патология почек, которая характеризуется поражением чашечек и лоханки почек (при анализе будет значительное увеличение количества лейкоциты, с нормальным или слегка измененным содержанием эритроцитов),
- Гломерулонефрит — аутоиммунная патология почек, при которой почечные клубочки поражаются циркулирующими иммунными комплексами (диагностические показатели будут противоположны пиелонефриту, т.е.е. увеличение количества эритроцитов будет выражаться нормальным или немного увеличенным количеством лейкоцитов).
- Цистит — воспалительное заболевание мочевого пузыря (количество лейкоцитов увеличивается, а количество эритроцитов остается неизменным)
- Гнойные процессы в почках (сопровождаются резким увеличением содержания лейкоцитов в моче и в норме для эритроцитов)
Причины возможных отклонений
Отклонениями принято считать расхождение с нормальными показателями более чем на 10%.Причины изменения количества форменных элементов в осадке мочи частично рассмотрены в предыдущем разделе. Чтобы понять причины изменения клеточного состава мочи, необходимо знать, за что отвечают эти клетки.
Лейкоциты — лейкоциты, основная функция которых — защита макроорганизма (в данном случае человека) от воздействия чужеродных агентов (органического и неорганического происхождения), они принимают участие во всех типичных воспалительных процессах (мигрируют из кровоток к очагу воспаления, постарайтесь впитать инородный агент и отграничить пораженный участок от здоровой ткани).
Следовательно, увеличение количества лейкоцитов в анализе по методу Амбурге дает понять, что где-то по длине от почек до уретры происходит воспалительный процесс. Точное место поражения определяется на основании жалоб, анамнеза и дополнительных методов исследования.
Эритроциты — это красные кровяные тельца, основная функция которых — перенос кислорода к органам и тканям. Отличительной особенностью эритроцитов является то, что они не стремятся покинуть кровоток в очаг поражения, а покидают его при нарушении структурной и функциональной целостности кровеносного сосуда.
Следовательно, если в результате анализа будет обнаружено повышенное содержание эритроцитов, то можно сказать, что где-то по длине от почек до уретры были повреждены кровеносные сосуды.
Чаще всего этому способствуют травмы слизистых оболочек мочевыводящих путей, мочекаменная болезнь, гломерулонефрит, опухолевый рост. Точную причину должен установить лечащий врач на основании жалоб, анамнеза и дополнительных методов исследования.
Detonic — уникальное лекарство, помогающее бороться с гипертонией на всех этапах ее развития.
Комплексное действие растительных компонентов препарата Детоник на стенки сосудов и вегетативную нервную систему способствует быстрому снижению артериального давления. Кроме того, этот препарат предотвращает развитие атеросклероза благодаря уникальным компонентам, которые участвуют в синтезе лецитина — аминокислоты, регулирующей метаболизм холестерина и предотвращающей образование атеросклеротических бляшек.
Детоник не вызывает привыкания и абстинентного синдрома, так как все компоненты продукта натуральные.
Подробная информация о Detonic находится на странице производителя www.detonicnd.com.
№ | Название услуги |
1 | Общий анализ крови (19 аппаратов) |
2 | Лейкоформула |
3 | Гемоглобин |
4 | Тромбоциты |
5 | Лейкоциты |
6 | Скорость оседания эритроцитов |
7 | Ретикулоцитов |
8 | Группа крови и резус-фактор |
9 | Перекрестное сопоставление |
10 | Тест Кумбса |
11 | Общий анализ мочи |
12 | Пигменты желчных камней в моче |
13 | Исследование осадка мочи под микроскопом |
14 | Общий анализ мочи методом Нечипоренко |
15 | Кетоны в моче |
16 | Белка в моче (в сутки) |
17 | Тест Амбурге |
18 | Проба Зимницкого |
19 | Анализ мочи по Каковски-Аддис метод |
20 | Тест на беременность |
21 | Общий тест стула |
22 | Анальный зуд |
23 | Анализ кала на скрытую кровь (FOBT) |
24 | Спермограмма (морфология, вид на мониторе) |
25 | Исследование жидкости предстательной железы |
26 | Микроскопия мазка из уретры |
27 | Микроскопия мазка из уретры (женский) |
28 | Микроскопия мазка из уретры (мужской и детский) |
29 | Chlamydia Trachomatis (иммунофлуоресцентный анализ (ИФА) |
30 | Уреаплазма в мазке из уретры |
31 | Микроплазма в мазке из уретры |
32 | Gardnerella vaginalis (одиночная) |
33 | Проба «Шуварского» (посткоитальная проба) |
34 | Тест Курцрока – Миллера |
35 | «Папоротник» проба |
36 | Демодекс на скине |
37 | Ресничные клещи или Demodex |
38 | Парша |
39 | Малярия (Густой мазок крови) |
40 | Общеклиническое исследование синовиальной жидкости |
41 | Исследование плеврального и перикардиального выпотов |
42 | Анализ перитонеальной жидкости |
43 | Анализ спинномозговой жидкости (ЦСЖ) |
44 | Исследование ила желчного пузыря |
Эрих Харнак (1852–1915) и краткая история апоморфина — FullText — European Neurology 2013, Vol.69, № 6
Аннотация
Апоморфин, в настоящее время признанный эффективным средством лечения рефрактерных двигательных колебаний при болезни Паркинсона, леченной леводопой, имеет долгую и неоднозначную историю применения в медицине и ветеринарии. Доклинические фармакологические эффекты апоморфина in vivo были впервые изучены около 150 лет назад, после чего препарат был введен для лечения поведенческих пороков у домашних животных.Ранние фармакологические исследования Эриха Гарнака в Дерпте (ныне Тарту, Эстония), где он принадлежал к фармакологической династии Буххаймов и Шмидебергов, имеют особое историческое значение, поскольку он подчеркивал, что, хотя апоморфин обладает сильным рвотным действием, препарат также оказывает комплексное воздействие на Центральная нервная система.
© 2013 S. Karger AG, Базель
Апоморфин, прототип агониста дофамина, который стимулирует рецепторы D1, D2 и D3, имеет долгую и интересную историю.Это будет кратко резюмировано, обращая особое внимание на малоизвестный вклад Эриха Харнака и Тартуской школы клинической фармакологии в Эстонии.
Осведомленность о влиянии апоморфина на мозг, вероятно, восходит к древним цивилизациям. Луковицы и корни кувшинки ( Nymphaea caerulea , голубой лотос и Nymphaea ampa с белым цветком), как теперь известно, содержат широкий спектр апорфинов, включая апоморфин.Ученые-этноботаники предположили, что видов Nymphaea использовались из-за своих наркотических, афродизиакальных и галлюциногенных свойств в магико-религиозных обрядах цивилизацией майя в Центральной Америке (2000 г. до н.э. — 250 г. н.э.). Их ритуальное использование также было изображено на фресках древних египетских гробниц и в ранних свитках папируса. Знаменитая древнеегипетская гробница Тутанхамона содержала золотую святыню, украшенную фараоном, держащим большую водяную лилию, а фрески в гробнице Небаума (XVIII династия, 1370–1318 гг. До н.э.) в Британском музее иллюстрируют погребальный танец с двумя женщинами, украшенными гирляндами. лепестки кувшинки [1].
Апоморфин был впервые синтезирован Арппе в 1845 году из морфина и серной кислоты ( Ueber eine merkwürdige Veränderung des Morphius durch Schwefelsäure . Liebig’s Annalen der Chemie und Pharmacie 1945; LV: 96), а позже введен Маттиезеном, представившимся Маттиезеном. синтез апоморфина из морфина и соляной кислоты (Proceedings of the Royal Society 1869; XVII: 455) [2]. Вскоре после открытия это лекарство было исследовано ветеринарами и рекомендовано для лечения стереотипов у домашних животных [3].
За первым комплексным фармакологическим исследованием эффектов апоморфина мы обратимся к Эриху Харнаку (рис. 1). Он родился в 1852 году в Дерпте, ныне Тарту, Эстония, в одаренной академической семье. Его отец, родившийся в Санкт-Петербурге, был профессором богословия, а его четыре сына, включая Эриха, выросли и стали профессорами фармакологии, теологии, физики и истории в различных европейских университетах [4,5]. В 1873 году Эрих Гарнак окончил Дерптский университет, один из старейших университетов Северной Европы, основанный в 1632 году, который по праву был назван «колыбелью современной фармакологии».Вскоре он присоединился к династии, основанной Рудольфом Буххаймом, который был главой отдела Materia Medica (фармакология) с 1847 по 1867 год и оказал влияние на раннее продвижение аналитической и экспериментальной работы в фармакологии, исходя из ее чисто описательных основ. : его экспериментальная фармакологическая лаборатория оставалась единственной в мире почти 20 лет. Это было развито его учеником Освальдом Шмидебергом, который, заняв кафедру, создал школу фармакологии, которая благодаря своей вдохновляющей учебной программе обеспечила руководителей многих академических институтов фармакологии по всему миру.Именно в этом интеллектуально стимулирующем месте в Старом анатомическом театре в Дерпте Гарнак начал свои исследования в области фармакологии животных (рис. 2) [6,7].
Рис. 1
Профессор Эрих Харнак (1852–1915).
Рис. 2
Старый анатомический театр Дерптского университета (ныне Тартуский университет, Эстония) Луи Хёфлингера (1860 г.).
Гарнак работал в Дерпте / Тарту в качестве доцента до 1877 года, затем переехал в Страсбург и, наконец, в университет Галле в качестве профессора фармакологии и физиологической химии, где в 1890 году основал новый Институт фармакологии [5].Вскоре он был признан выдающимся ученым-новатором и популярным учителем. Однако его эксцентричность и либерализм не всегда ценились негибкой академической иерархией. Иногда он читал лекции вне университета в кафе, где студентам разрешалось курить и пить пиво [6,7]. Он поощрял и смаковал критику, особенно когда она была окрашена нежной сатирой. В статье The New York Times за 1909 год он высмеивал «безалкогольное пиво» и «кофе без кофеина» как «ослабленные стимуляторы» и «самообман».У него было захватывающее причудливое чувство юмора, и он считал жизнь с сопутствующими ей рисками, опасностями и искушениями предпочтительнее, чем жизнь в искусственно созданном «идеальном» безопасном сообществе [8].
Харнак широко публиковал статьи в области фармакологии, в том числе статьи о действии алкоголя и симптомах отравления картофелем, а также заинтересовался медицинским использованием электричества и магнетизма. Он написал учебники по медицине, а его докторская диссертация была посвящена патогенезу и лечению сахарного диабета [9].
В 1874 году в возрасте 22 лет, работая в Дерпте, Эрих Харнак опубликовал 50-страничный отчет « Ueber die Wirkungen des Apomorphins am Saugerthier und am Frosch » (Эффекты апоморфина у млекопитающих и лягушек) в журнал « Archiv für Experimentelle Pathologie und Pharmakologie », основанный в 1873 году, ныне известный как Архив фармакологии Наунин-Шмидеберг [10]. К тому времени его коллега Винсент Зиберт, также из Дерптского университета (Тарту), уже изучил рвотные эффекты апоморфина и установил, что связанные с ним изменения кровообращения ограничены определенными видами, включая людей, собак и кошек, но не лягушек или кроликов; но в своей диссертации 1871 года Зиберт, которого руководил Освальд Шмидеберг, пришел к выводу, что знания о возможных эффектах апоморфина на центральную нервную систему «остаются окутанными тьмой» [2].
Во введении Харнак подчеркнул, что, хотя наркотики могут иметь очевидные эффекты, такие как рвота, могут быть и другие менее очевидные, но важные действия, в равной степени заслуживающие внимания, и именно этот принцип стал его основной экспериментальной стратегией в отношении апоморфина. Помимо подробного обзора литературы, Харнак описал свои собственные обширные эксперименты на животных. К ним относятся исследования воздействия апоморфина в различных дозах на собак, кошек, кроликов и лягушек.Он обратил особое внимание на тот факт, что рвотные дозы апоморфина у детей могут одновременно вызывать коллапс кровообращения, а у взрослых дозы 5–10 мг только вызывают рвоту. Уже было известно, что более высокие дозы вызывают двигательные эффекты у кошек и собак, и чтобы прояснить эти различия в действиях, Харнак изучал кроликов, у которых не было рвотной реакции. После парентерального введения 2 мг апоморфина было спровоцировано возбужденное двигательное поведение, которое усиливалось после повторных доз.При более высоких дозах животные непрерывно облизывали и грызли подстилку и стенки клеток. Он отметил, что кумулятивные дозы 10 мг вызывают паралич мышц, дыхательную недостаточность и терминальные судороги. Дальнейшие исследования показали, что судороги были вызваны не гипоксией из-за дыхательной недостаточности, а раздражением центральной нервной системы. Хлороформная анестезия подавляла рвоту, вызванную апоморфином, но не влияла на дыхание, указывая на отдельные центральные центры дыхания и рвоты.В своих исследованиях на лягушках Гарнак заметил, что за двигательной стимуляцией следует паралич мышц. Он пришел к выводу, что низкие дозы стимулируют центр рвоты, а большие — его подавляют; у кроликов без рвотного ответа дыхательный центр стимулировался более низкими дозами и подавлялся более высокими дозами; у собак наблюдались только стимулирующие эффекты. Он также наблюдал различия этих эффектов при сравнении апоморфина и морфина: центральные стимулирующие эффекты были более выраженными у апоморфина, но после морфина стадия ингибирования начиналась быстрее и была более выраженной, чем после апоморфина [10].
Его эксперименты привели его к выводу, что апоморфин действует в нескольких различных центрах мозга, включая те, которые участвуют в произвольном поведении, сердечно-сосудистом и дыхательном контроле и ощущениях. Эта работа была проведена почти на столетие раньше, чем исследования Andén et al. (1967) и Эрнст (1967), которые впервые продемонстрировали, что апоморфин напрямую активирует специфические рецепторы дофамина и что нейролептики блокируют рецепторы дофамина и действуют как антагонисты апоморфина [11,12].
Примерно в то же время, когда Гарнак проводил свои фармакологические исследования, Сэмюэл Джи в Лондоне изучал рвотные эффекты апоморфина в низких дозах у людей, демонстрируя его значительный потенциал как рвотного средства [13]. Самое раннее упоминание о потенциальном использовании апоморфина для лечения болезни Паркинсона было сделано Вейлем в 1884 году, хотя к тому времени он был предложен для лечения танца Святого Вита [14].
В течение 20-го века было проведено множество подробных исследований многочисленных моторных и поведенческих эффектов, включая зевание и эрекцию полового члена, что привело к идентификации различных типов дофаминовых рецепторов для объяснения разнообразных клинических и экспериментальных последствий введения апоморфина.Введение блокаторов дофаминовых рецепторов значительно облегчило понимание различных эффектов апоморфина, а также его использование в клинической практике [15,16]. Эффекты апоморфина в дальнейшем использовались, часто спорно, для самых разных расстройств, от лечения гомосексуализма, алкогольной, табачной и морфиновой зависимости до эректильной дисфункции, но это были бостонские невролог Роберт Шваб (1951) и клинический ученый Джордж Котзиас. (1970), которые окончательно подтвердили предположение Вейля о том, что апоморфин может иметь антипаркинсонический эффект [17,18].Оценка современного использования апоморфина в оценке, диагностике и лечении моторных осложнений болезни Паркинсона выходит за рамки данного отчета [19].
Решающий вклад Эриха Гарнака состоял в том, чтобы продемонстрировать сильное влияние апоморфина на центральную нервную систему. Его это позабавило бы, если бы не удивило, что после демонстрации его эффективности при болезни Паркинсона ряд регулирующих лекарств органов отложили повторное введение апоморфина в современную терапию из-за необоснованных опасений, связанных с его химическим сходством с морфином.
Благодарности
Мы благодарим коллег из библиотеки Тартуского университета, кафедры фармакологии и Старого анатомического театра Тартуского университета, а также из университетского архива Университета Мартина Лютера в Галле-Виттенберге.
Заявление о раскрытии информации
У авторов нет конфликта интересов, о котором следует раскрывать.
Список литературы
- Бертоль Э, Финески В., Карч С.Б., Мари Ф., Риеццо I: Нимфеи культов в Древнем Египте и Новом Свете: урок эмпирической фармакологии.JR Soc Med 2004; 97: 84–85.
- Зиберт V: Untersuchungen über die Physiologischen Wirkungen des Apomorphin. Вступительная диссертация zur Erlangung der Doctorgrades. Дерпт, Друк фон Генрих Лаакманн, 1871 г.
- Фесер Дж .: Умереть в neuester Zeit в Anwendung gekommen Arzneimittel: 1.Апоморфин гидрохлорат. Z Prakt Veterinairwiss 1873: 302–306.
- Национальный архив Эстонии: архивы EAA.402.2.8802–8812, 1824–1877.
- Welding O, Amburger E, von Krusenstjern G, Lenz W. (ред.): Deutschbaltisches Biographisches Lexicon.Кёльн-Вена, Böhlau Verlag, 1970.
- Тоомсалу М: профессора Старого анатомического театра Тартуского университета. Издательство Тартуского университета, 2006.
- Кущинский К. Антология из архива фармакологии Наунин-Шмидеберг: E.Гарнак (1874 г.): Ueber die Wirkungen des Apomorphins am Säugethier und am Frosch. Archiv für Experimentelle Pathologie und Pharmakologie 2: 254–306 (О влиянии апоморфина на млекопитающих и лягушек). Naunyn Schmiedebergs Arch Pharmacol 2006; 373: 387–389.
- Harnack E: Критикует названия напитков умеренности.Нью-Йорк Таймс 1909 г .; 11.
- Harnack E: Zur Pathogenese und Therapie des Diabetes mellitus. Вступительная диссертация zur Erlangung des Grades eines Doctors der Medicin. Дорпат, Gedruckt bei Heinrich Laakmann, 1873.
- Harnack E: Ueber die Wirkungen des Apomorphins am Säugethier und am Frosch.Arch Exp Pathol Pharmakol 1874; 2: 254–306.
- Andén NE, Rubenson A, Fuxe K, Hökfelt T: Доказательства стимуляции дофаминовых рецепторов апоморфином. J Pharm Pharmacol 1967; 19: 627–629.
- Эрнст AM: Механизм действия апоморфина и дексамфетамина на принуждение к грызу у крыс.Psychopharmacologia 1967; 10: 316–23.
- Джи С: О действии новой органической основы, апоморфии. Trans Clin Soc Lond 1869; 2: 166–169.
- Вейл Э: Апоморфин вызывает определенные нервные расстройства.Lyon Méd 1884; 48: 411–419.
- Штерн G, Аполавстический апоморфин. Pract Neurol 2013 (в печати).
- Серра Г., Фратта В., Коллу М., Наполи-Фаррис Л., Гесса Г.Л.: Циклогексимид предотвращает вызванное апоморфином зевание, эрекцию полового члена и уход за гениталиями у крыс.Eur J Pharmacol 1982; 86: 279–282.
- Schwab RS, Amador LV, Lettvin JY: Апоморфин при болезни Паркинсона. Trans Am Neurol Assoc 1951; 56: 251–253.
- Cotzias GC, Papavasilou PS, Kaufman B, Mena I. Сходства между неврологическими эффектами L-допа и апоморфина.N Engl J Med 1970; 282: 31–33.
- Стиб CMH, Лис А.Дж., Кемпстер П.А., Стерн Г.М.: Подкожный апоморфин в паркинсонических колебаниях включения-выключения. Ланцет 1988; 1: 403–406.
Автор Контакты
Пилле Таба, доктор медицинских наук
Кафедра неврологии и нейрохирургии
Тартуский университет, Пуусепа 8 — h552
EE – 51014 Тарту (Эстония)
E-Mail [email protected]
Подробности статьи / публикации
Предварительный просмотр первой страницы
Получено: 30 октября 2012 г.
Принято: 24 декабря 2012 г.
Опубликовано в Интернете: 14 марта 2013 г.
Дата выпуска: июнь 2013 г.
Количество страниц для печати: 4
Количество рисунков: 2
Количество столов: 0
ISSN: 0014-3022 (печатный)
eISSN: 1421-9913 (онлайн)
Для дополнительной информации: https: // www.karger.com/ENE
Авторские права / Дозировка препарата / Заявление об ограничении ответственности
Авторские права: Все права защищены. Никакая часть данной публикации не может быть переведена на другие языки, воспроизведена или использована в любой форме или любыми средствами, электронными или механическими, включая фотокопирование, запись, микрокопирование, или с помощью любой системы хранения и поиска информации, без письменного разрешения издателя. .
Дозировка лекарства: авторы и издатель приложили все усилия, чтобы гарантировать, что выбор и дозировка лекарства, указанные в этом тексте, соответствуют текущим рекомендациям и практике на момент публикации.Однако с учетом продолжающихся исследований, изменений в правительственных постановлениях и постоянного потока информации, касающейся лекарственной терапии и реакций на них, читателю рекомендуется проверять листок-вкладыш для каждого препарата на предмет любых изменений показаний и дозировки, а также дополнительных предупреждений и меры предосторожности. Это особенно важно, когда рекомендованным агентом является новое и / или редко применяемое лекарство.
Отказ от ответственности: утверждения, мнения и данные, содержащиеся в этой публикации, принадлежат исключительно отдельным авторам и соавторам, а не издателям и редакторам.Появление в публикации рекламы и / или ссылок на продукты не является гарантией, одобрением или одобрением рекламируемых продуктов или услуг или их эффективности, качества или безопасности. Издатель и редактор (-ы) не несут ответственности за любой ущерб, причиненный людям или имуществу в результате любых идей, методов, инструкций или продуктов, упомянутых в контенте или рекламе.
Amburger (США) комментарии к The Experiment Отображение 1-20 из 725
Я просто понял, что никогда не был участником этого конкретного рп.Я был в другой группе. Ха-ха.Ага. Финалы важны. Сосредоточьтесь на них, ха-ха. Я тоже очень боюсь своего.
Это так много. Можете ли вы подвести итог для …. других? кто ленивый?
Ничего страшного. он сказал, что может быть свободен в эти выходные. Мощь. Не знаю, откажусь ли я от него или что-то в этом роде. но МЫ увидим.
Я имею в виду, я никогда не участвовал в этом. Итак … Я не буду помнить, что происходит, ха.
Я не знаю. Мы собирались встретиться в это воскресенье перед тем, как провести время в субботу. Суббота была как бы последней минутой, потому что его репетиция была отменена. Так что я спрошу, например, в четверг, хочет ли он по-прежнему проводить время в воскресенье. Хотя я не уверен.
Если людей не так много (а их трое — не много), я бы сказал, что нет? Если только вы не хотите его только для целей организации или чего-то в этом роде.
Он танцует хип-хоп, ха-ха. Я не могу танцевать.
YEEEEE. достать их, мальчики. Думаю, вчера вечером я вроде как пошла на свидание…. я чувствую тебя
Ух ты. Не думаю, что когда-либо участвовал в этой ролевой игре. Но … в эти выходные у тебя было еще одно двойное свидание. Как это было? (Мне очень жаль, что я вернулся к теме свиданий)
Ооооо. Как было свидание ?? Я имею в виду, я думаю, что если вы полностью в твиттере, это было бы хорошо, но подробнее.
это хорошо, это хорошо. Так как дела? Я правда думал, что ладил чаще, чем я, но, видимо, нет.
Ух ты.Ладно, по-видимому, я больше не занимаюсь этим часто … Я не занимаюсь … сочинительством весело, без ролевых игр, ха-ха.
Это было навсегда. Я обычно проверяю примерно … раз в неделю, если мне хочется. Если я получаю хорошие чтения, когда пытаюсь ввести текст на других веб-сайтах.
((Я все еще тут и выключаюсь. Я вижу, что ты вернулся, ха-ха.))
О боже. Почему? Если хотите, поделитесь.
Не думаю, что кто-нибудь вспомнит. Так давно.
«Что …. Что я сделал?» — спросила Хана, оборачиваясь и глядя на огонь, который она развела. «Да ладно. Он может получить новую дверь. Нам нужно место, чтобы остаться».
Хана кивнула. «Грубый придурок». пробормотала она. В гневе она не осознавала, что оставила на двери табличку. Как только они отошли, дверь дома загорелась.
Хана на секунду посмотрела на мальчика, задумавшись. «На улице холодно». Наконец она сказала отрывисто.Она начала протискиваться мимо мальчика, но потом поняла, что это может показаться грубым. Она остановилась и отступила на несколько шагов, позволяя Эш взять под контроль.
((Каждый раз, когда я слышу Эша … я представляю … Эша … в Лиге Легенд.. … как … это все, что я представляю.))
Pulmonary Daoyin как программа реабилитации пациентов с ИЛФ в традиционной китайской медицине: рандомизированное контролируемое исследование
Респирология. 2021 Apr; 26 (4): 360–369.
, 1 , 2 , * , 1 , 3 , * , 2 , 4 , 5 , 6 , 7 и 1 , 8Мяо Чжоу
1 Совместное строительство Совместного инновационного центра китайской медицины и респираторных заболеваний при Хэнань и Министерстве образования П.Р. Китай, Хэнаньский университет китайской медицины, Чжэнчжоу Китай,
2 Отделение респираторных заболеваний, Третья дочерняя больница Хэнаньского университета китайской медицины, Чжэнчжоу Китай,
Хайлун Чжан
1 Совместное строительство инновационного центра китайской медицины и респираторных заболеваний при Министерстве образования Китая и провинции Хэнань, Хэнаньский университет китайской медицины, Чжэнчжоу Китай,
3 Отделение респираторных заболеваний, Первая дочерняя больница Хэнаньского университета китайской медицины, Чжэнчжоу Китай,
Фенглей Ли
2 Отделение респираторных заболеваний, Третья дочерняя больница Хэнаньского университета китайской медицины, Чжэнчжоу Китай,
Чжэфэн Юй
4 Отделение респираторных заболеваний, Больница китайской медицины Ручжоу, Пиндиншань Китай,
Чэнбо Юань
5 Отделение респираторных заболеваний, Аффилированная больница Чанчуньского университета китайской медицины, Чанчунь Китай,
Брайан Оливер
6 Школа естественных наук, факультет естественных наук, Сиднейский технологический университет, Сидней Новый Южный Уэльс, Австралия,
7 Институт медицинских исследований Вулкока, Сиднейский университет, Сидней Новый Южный Уэльс, Австралия,
Цзяньшэн Ли
1 Совместное строительство Совместного инновационного центра китайской медицины и респираторных заболеваний при Хэнань и Министерстве образования П.Р. Китай, Хэнаньский университет китайской медицины, Чжэнчжоу Китай,
8 Хэнань ключевая лаборатория китайской медицины респираторных заболеваний, Хэнаньский университет китайской медицины, Чжэнчжоу Китай,
1 Совместное строительство инновационного центра китайской медицины и респираторных заболеваний при Министерстве образования Китая и провинции Хэнань, Хэнаньский университет китайской медицины, Чжэнчжоу Китай,
2 Отделение респираторных заболеваний, Третья дочерняя больница Хэнаньского университета китайской медицины, Чжэнчжоу Китай,
3 Отделение респираторных заболеваний, Первая дочерняя больница Хэнаньского университета китайской медицины, Чжэнчжоу Китай,
4 Отделение респираторных заболеваний, Больница китайской медицины Ручжоу, Пиндиншань Китай,
5 Отделение респираторных заболеваний, Аффилированная больница Чанчуньского университета китайской медицины, Чанчунь Китай,
6 Школа естественных наук, факультет естественных наук, Сиднейский технологический университет, Сидней Новый Южный Уэльс, Австралия,
7 Институт медицинских исследований Вулкока, Сиднейский университет, Сидней Новый Южный Уэльс, Австралия,
8 Хэнань ключевая лаборатория китайской медицины респираторных заболеваний, Хэнаньский университет китайской медицины, Чжэнчжоу Китай,
Автор, ответственный за переписку. * Для переписки: Цзяньшэн Ли, Совместное строительство инновационного центра китайской медицины и респираторных заболеваний при Хэнане и Министерстве образования КНР, Хэнаньский университет китайской медицины, Университетский городок Лунцзиху, Новый район Чжэндун, Чжэнчжоу, Хэнань 450046, Китай. Эл. Почта: moc.361@8sj_il,* M.Z. и Х.З. внесли равный вклад в это исследование.
Поступило 10.02.2020 г .; Пересмотрено 30 сентября 2020 г .; Принято 20 октября 2020 г.
Copyright © 2020 Авторы. Respirology , опубликованная John Wiley & Sons Australia, Ltd от имени Азиатско-Тихоокеанского общества респирологии. Это статья в открытом доступе на условиях http://creativecommons.org/licenses/by-nc-nd/4.0/ Лицензия, которая разрешает использование и распространение на любом носителе при условии правильного цитирования оригинальной работы, использования в некоммерческих целях и без внесения изменений или адаптаций. Эта статья цитируется в других статьях PMC.- Заявление о доступности данных
Индивидуальные данные участников, которые лежат в основе результатов, представленных в этой статье, и протокол исследования могут быть доступны после деидентификации (текст, таблицы, рисунки и приложения) от 9 до 36 месяцев после публикации статьи для отдельных лиц. метаанализ данных участников.Предложения исследователя об использовании данных должны быть одобрены независимым комитетом по анализу, назначенным для этой цели с подписанным соглашением о доступе к данным.
РЕФЕРАТ
Предпосылки и цель
IPF — это хроническое прогрессирующее заболевание легких, при котором PR приносит пользу пациентам. PD, программа PR TCM, известна своей эффективностью при ХОБЛ, но ее полезность при IPF неизвестна. Мы исследовали его эффективность и безопасность у пациентов с ИЛФ.
Методы
Шестимесячное рандомизированное контролируемое исследование (РКИ) было проведено в трех китайских клиниках.Девяносто шесть участников с диагнозом IPF были случайным образом распределены в одну из трех групп: группа PD получала программу PD два раза в день, 5 дней в неделю в течение 2 месяцев, а группа упражнений выполняла упражнения на стационарном велоэргометре, 30 мин. / день, 5 дней / неделя в течение 2 месяцев. Добровольцам контрольной группы посоветовали продолжать свою обычную деятельность. Первичные исходы заключались в изменении по сравнению с исходным уровнем показателей 6MWD и HRQoL по SGRQ-I через 1 и 2 месяца (в конце вмешательства) и через 6 месяцев (через 4 месяца после вмешательства).Вторичные показатели исходов включали FVC, DL CO (прогнозируемый%) и изменения в mMRC.
Результаты
6MWD был увеличен в группе PD по сравнению с группами упражнений и контрольной группой. 6MWD увеличился на 60,44 м в группе PD, 32,16 м в группе упражнений и 12,42 м в контрольной группе после 2 месяцев реабилитационной программы. Межгрупповые различия в изменении от исходного уровня составили 28,78 м (95% ДИ: от 0,54 до 56,01; P = 0,044) и 48,02 м (95% ДИ: 23.С 04 до 73.00; P <0,001) через 2 месяца и 25,61 м (95% ДИ: от -0,67 до 51,89; P = 0,058) и 50,93 м (95% ДИ: от 25,47 до 76,40; P <0,001) через 6 месяцев соответственно, включая разницу, превышающую MCID. Не было значительных изменений в оценке SGRQ-I, оценке одышки mMRC, FVC и DL CO (% от прогнозируемого) ни в группах PD, ни в группах упражнений.
Заключение
Через два месяца после вмешательства наблюдалась клинически значимая разница в 6MWD в пользу программы PD.Программа PD безопасна и эффективна в качестве реабилитационного вмешательства, направленного на повышение толерантности к физической нагрузке, и является подходящей заменой PR.
Ключевые слова: 6 минут ходьбы, Даоинь, идиопатический фиброз легких, легочная реабилитация, традиционная китайская медицина
Сокращения
- 6MWD
- 6 минут ходьбы
- AE
- 92 неблагоприятное событие IP
- острое обострение IPF
- ANOVA
- дисперсионный анализ
- ATS / ERS / JRS / ALAT
- Американское торакальное общество / Европейское респираторное общество / Японское респираторное общество / Латиноамериканская торакальная ассоциация
- Хроническая обструктивная болезнь легких ХОБЛ
- DL CO
- диффузионная способность легких по монооксиду углерода
- FVC
- форсированная жизненная емкость
- HRQoL
- качество жизни, связанное со здоровьем
- IPF
- идиопатический легочный фиброз 908-9 лечить
- MCID
- минимально клинически важное различие
- mMRC
- модифицированный Британский совет медицинских исследований
- PD
- легочный Daoyin
- PR
- легочная реабилитация
- RCT
- рандомизированное контролируемое исследование
- SGRQ ‐ I
- St George’s 1 — T 0
- изменения в конечной точке первого месяца исследования по сравнению с исходным уровнем
- T 2 — T 0
- изменения в конце реабилитационного исследования по сравнению с исходным уровнем
- T 6 — T 0
- изменения в конце контрольного исследования по сравнению с исходным уровнем
- TCM
- традиционная китайская медицина
ВВЕДЕНИЕ
Идиопатический фиброз легких (IPF) — хроническое заболевание с переменным течением болезни, при котором прогрессирует происходит ухудшение функции легких, что приводит к значительной заболеваемости и m ортодоксальность.Средняя выживаемость составляет 2–3 года. 1 Заболеваемость составляет 2–30 случаев на 100 000 человеко-лет, а распространенность колеблется от 10 до 60 случаев на 100 000 человек. 2 , 3 , 4 , 5 , 6 в зависимости от исследуемой страны. Эти цифры примерно соответствуют численности населения 130 000 в Соединенных Штатах, 300 000 в Европе, 640 000 в Восточной Азии и ~ 3 миллиона человек во всем мире. 7 У пациентов в возрасте> 65 лет предполагаемая распространенность ИЛФ достигает 400 случаев на 100 000 человек в США.8 Данных о распространенности из Китая мало, но текущие данные показывают, что заболеваемость ИЛФ увеличивается. Увеличение количества госпитализаций и смертей из-за ИЛФ также свидетельствует об увеличении бремени болезней. 9
За последние 5 лет были достигнуты заметные успехи в фармакотерапевтическом лечении ИЛФ с появлением нинтеданиба и пирфенидона. Оба они безопасны и эффективны при лечении пациентов с ИЛФ. 10 , 11 Однако отсутствие продолжительного времени выживания и их высокая стоимость ограничивают их глобальное клиническое применение.Стратегии немедикаментозного лечения снижают заболеваемость, и важность этих подходов невозможно переоценить. 9 Дополнительная кислородная терапия и легочная реабилитация (PR) настоятельно рекомендуются пациентам с IPF в клинических рекомендациях. 11 , 12 PR, структурированная программа упражнений, разработанная для взрослых с тяжелыми заболеваниями легких, 9 было обнаружено, что он улучшает расстояние ходьбы, симптомы и качество жизни пациентов с ИЛФ. 13 , 14 , 15
Легочный Даоинь (PD), PR-технология традиционной китайской медицины (TCM), была создана на основе навыков Даоинь.16 Хотя эффекты БП у пациентов с хронической обструктивной болезнью легких (ХОБЛ) были описаны в предыдущем исследовании, 17 Эффективность программы у пациентов с ИЛФ неизвестна. В настоящем исследовании изучалась эффективность и безопасность ПД по сравнению с обычным уходом у пациентов с ИЛФ.
МЕТОДЫ
Дизайн исследования
Это исследование было рандомизированным клиническим исследованием. Субъекты были случайным образом распределены в одну из трех групп: PD, упражнения и контрольная группа.Случайное распределение было выполнено с использованием программного обеспечения рандомизатора (SAS 9.4). Исследование было зарегистрировано в Китайском реестре клинических испытаний (ChiCTR ‐ IOR ‐ 17011187) 19 апреля 2017 г. Этическое одобрение было получено Комитетом по этическим исследованиям Первой аффилированной больницы Хэнаньского университета китайской медицины (номер партии — 2016HL ‐ 083) ). Письменное информированное согласие было получено от всех участников.
Размер выборки
Размер выборки был основан на влиянии PR на 6-минутное расстояние ходьбы (6MWD) в IPF.Джексон и др. . 18 обнаружили, что PR в течение 3 месяцев улучшил 6MWD пациентов IPF (354,4 ± 37,2 против 324,1 ± 35,5) с уровнем значимости 5%, мощностью 0,90 при уровне значимости 5% (двусторонний), следовательно, 32 субъекта на группа была обязательна.
Объекты исследования
Субъектам был поставлен диагноз IPF в соответствии с диагнозом IPF (официальное руководство по клинической практике Американского торакального общества / Европейского респираторного общества / Японского респираторного общества / Латиноамериканской торакальной ассоциации (ATS / ERS / JRS / ALAT)).11 Критерии включения включали стабильный IPF и возраст от 40 до 85 лет. Критериями исключения были сатурация кислорода в покое <90%, когнитивные нарушения / психические заболевания, обмороки после упражнений, остеоартрит и тяжелые сопутствующие заболевания, включая острый инфаркт миокарда и нестабильную стенокардию.
Протокол вмешательства
ПД группаPD, технология TCM PR, была создана на основе навыков Даоинь и теории TCM для пациентов с ХОБЛ.16 Это мягкая медитативная техника, которая применяет физические движения, дыхательные упражнения и регуляцию ума. Он основан на принципе интеграции и гармонизации ума, дыхания, позы и движения, включая тренировку дыхательных мышц. Подробные шаги и графическое описание позы программы PD можно найти в протоколе исследования, опубликованном нашей командой. 16
Субъекты в группе PD завершили двухмесячную программу PD, которая состояла из двух этапов.Первый этап — это месячная фаза интенсивных упражнений. На этом этапе испытуемые тренировались 5 дней в неделю. Им разрешили разделить 1-часовую тренировку на утренние и дневные занятия. Класс PD проводил квалифицированный инструктор PD. Методы PD были повторно оценены через 1 месяц, чтобы убедиться в сохранении надлежащих навыков. Второй этап представлял собой месячную фазу физических упражнений без присмотра дома 5 дней в неделю, два раза в день. Наряду с письменными инструкциями каждому испытуемому был выдан компакт-диск, чтобы облегчить ежедневную самостоятельную практику.Каждому испытуемому также был предоставлен дневник для записи частоты их сеансов.
Группа упражненийТренировки с упражнениями представляли собой общую программу (не специфичную для IPF), которая включала амбулаторную программу тренировок пять раз в неделю, интегрированную с тренировкой периферических мышц. Перед началом программы тренировок с физической нагрузкой измеряли артериальное давление, сатурацию кислорода и частоту сердечных сокращений, чтобы оценить потенциальную потребность в оксигенации и обеспечить безопасность вмешательства.Целевая частота пульса и интенсивность упражнений определялись по результатам теста с 6-минутной ходьбой. Программа упражнений состояла из 5 минут разминки мышц и сердечно-сосудистой системы испытуемого перед тренировкой на стационарном велоэргометре. После достижения целевой частоты пульса тренировка продолжалась с такой же интенсивностью в течение 20 минут. Артериальное давление, сатурация кислорода и частота сердечных сокращений были снова измерены после тренировки. Наконец, была проведена 10-минутная тренировка релаксации.Дополнительный кислород подавался для поддержания насыщения кислородом выше 90%, если наблюдалась десатурация. Испытуемые в группе упражнений выполнили двухмесячную программу упражнений.
Контрольная группаСубъектам контрольной группы было рекомендовано сохранять свою обычную активность. Никаких дополнительных упражнений не рекомендовалось.
Оценка
Первичными исходами были 6MWD и респираторный опросник Святого Георгия для IPF (SGRQ-I). 6MWD проводился в соответствии с критериями ATS.19 SGRQ-I — полезный инструмент для измерения качества жизни, связанного со здоровьем (HRQoL) у пациентов с IPF, который может использоваться у пациентов с различной продолжительностью заболевания. 20 Он оценивает влияние респираторных заболеваний по трем параметрам: симптомы, активность и психосоциальное воздействие, а также дает общую оценку. Чем выше балл, тем тяжелее бремя IPF для человека.
Вторичными исходами были форсированная жизненная емкость легких (FVC), диффузионная способность легких по монооксиду углерода (DL CO ) и модифицированная шкала одышки Британского совета медицинских исследований (mMRC).Функция легких, FVC и DL CO коррелируют с исходами болезни IPF, с более серьезными нарушениями, связанными со снижением HRQoL 21 год и выживание. 22 Шкала mMRC — это инструмент самооценки, позволяющий измерить степень инвалидности, которую вызывает одышка при повседневной деятельности, по шкале от 0 (одышка отсутствует, кроме тяжелых физических упражнений) до 4 (слишком одышка, чтобы выйти из дома, или одышка. при одевании или раздевании). 23 , 24 Увеличение показателя mMRC отражает ухудшение качества жизни сердца и большое количество симптомов.Он использовался в качестве простого инструмента скрининга потребностей пациентов с ИЛФ в паллиативной помощи. 25 и один из показателей оценки эффективности в клинических испытаниях, связанных с IPF. 26 год
Сбор данных по 6MWD, SGRQ-I и mMRC проводился на исходном уровне, в месяцы 1 и 2 (конец вмешательства) и через 6 месяцев (через 4 месяца после вмешательства). Основными исходами были изменения по сравнению с исходным уровнем в 6MWD и SGRQ-I через 1 и 2 месяца вмешательства и через 6 месяцев. Результаты FVC и DL CO регистрировали на исходном уровне и через 2 месяца.
Статистический анализ
Анализ данных проводился с использованием программного обеспечения SAS 9.2 (SAS Institute Inc., Кэри, Северная Каролина, США). Описательная статистика использовалась для определения демографических характеристик выборки. Односторонний дисперсионный анализ (ANOVA) использовался для сравнения средних групп. Парные t-тесты (с 95% доверительным интервалом) использовались для изучения изменений внутри группы от исходного уровня до 1, 2 и 6 месяцев. В качестве уровня значимости было принято значение P 0,05 (двусторонний). Чтобы сохранить ценность рандомизации, мы использовали анализ намерения лечить (ITT).Отсутствующие данные об исключенных участниках были заменены методом переноса последнего наблюдения.
РЕЗУЛЬТАТЫ
Участники и исходные характеристики
С апреля 2017 года по август 2018 года было набрано в общей сложности 169 пациентов (83 были набраны из Третьей дочерней больницы Хэнаньского университета китайской медицины, 41 из больницы китайской медицины Ручжоу и 45 из Аффилированной больницы Чанчуньского университета китайской медицины) из амбулаторного отделения и проверены на соответствие критериям.В общей сложности 96 субъектов были случайным образом распределены в каждую из следующих групп: группа PD ( n = 32), группа упражнений ( n = 31) и контрольная группа ( n = 33). 2 месяца реабилитации прошли 92 пациента (95,8%); 89 пациентов (92,7%) завершили исследование. Все рандомизированные пациенты были включены в популяцию ITT (рис.). Демографические и исходные характеристики представлены в таблице. Не было значительных различий в клинически значимых исходных характеристиках между тремя группами.
Блок-схема исследования для отслеживания участников в ходе рандомизированного контролируемого исследования. AE ‐ IPF, обострение идиопатического фиброза легких; ITT, намерение лечить; PD, легочный Даоинь.
Таблица 1
Демографические и клинические характеристики участников исследования на исходном уровне
| Контрольная группа ( n = 31) | Группа упражнений ( n = 31) | PD группа ( n = 32) | ANOVA P -value | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Возраст, лет, среднее значение (SD) | 67 ± 10 | 64 ± 9 | 66 ± 11 | 0.497 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Пол | 0,965 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Муж. ) | 14 (45,16) | 13 (41,94) | 13 (40,62) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Рост, см, среднее (стандартное отклонение) | 168,19 ± 6,99 | 167,33 ± 6,28 | 167,22 0,811 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Масса, кг, средняя (стандартное отклонение) | 67.19 ± 12,53 | 65,85 ± 13,30 | 65,97 ± 9,27 | 0,884 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Сопутствующие заболевания | 0,269 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 1246 911 411 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 1246 911 (12,90) | 2 (6,25) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Ишемическая болезнь сердца (%) | 3 (9,68) | 0 | 1 (3,125) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Диабет | (%) 1 9012 3.23) | 0 | 2 (6,25) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Аллергический ринит (%) | 0 | 0 | 1 (3,125) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| гиперплазия простаты | % 01 (3,125) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| mMRC, среднее (SD) | 2,77 ± 0,99 | 2,77 ± 1,15 | 2,81 ± 1,12 | 0,987 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 6MWD, среднее | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 6MWD 281,52 ± 106,84 | 279.10 ± 108,08 | 284,06 ± 129,20 | 0,986 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ФЖЕЛ, L, среднее (стандартное отклонение) | 2,12 ± 0,49 | 2,25 ± 0,70 | 2,17 ± 0,49 | 0,667 | 0,667 | 90V119 L, среднее (стандартное отклонение)1,54 ± 0,48 | 1,67 ± 0,57 | 1,54 ± 0,46 | 0,478 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DL CO (% от прогноза) | 62,43 ± 7,80 | 46 65,17 ± 14,6046 65,17 ± 14,60 9,790.492 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BBS, среднее (стандартное отклонение) | 42,73 ± 13,78 | 45,17 ± 9,17 | 42,43 ± 14,69 | 0,663 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| SGRQ-I (симптомы) | ± 1511684,8 | 84,93 ± 18,96 | 0,996 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| SGRQ-I (Активность) | 80,05 ± 26,27 | 79,43 ± 20,69 | 80,36 ± 21,83 | 0,987 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 76,22 ± 23,17 | 74,98 ± 26,87 | 0,974 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| SGRQ ‐ I (Всего) | 78,94 ± 24,83 | 78,66 ± 19,39 | 78,39 ± 23,063 | 9115|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Измерение | Контроль | Упражнение | PD | Разница между группами в изменении от исходного уровня | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ( n = 31) | ( n = 32) | PD против упражнения | PD против контроля | Упражнение против контроля | |||||||||||||||
| P -значение | 95% ДИ | P -значение | 95% ДИ | P -значение | 95% ДИ | P -значение | |||||||||||||
| 6MWD (м) | 9 | ||||||||||||||||||
| 9 7.55 ± 16,00 | 13,55 ± 24,42 | 28,88 ± 36,51 | 0,007 | 15,33 (от -3,88 до 34,53) | 0,154 | 21,33 (от 3,80 до 38,86) | 0,013 | -611,9 0,586||||||||||||
| T 2 — T 0 | 12,42 ± 21,20 | 32,16 ± 35,45 | 60,44 ± 52,63 | 0,001 | 28,78 (от 0,54 до 56,01 9114 0125 | от 0,54 до 56,01 9125 | )0.001 | 19,74 (от 1,43 до 38,06) | 0,031 | ||||||||||
| T 6 — T 0 | 3,16 ± 36,54 | 28,48 ± 39,45 | 5441146 25,67 до | 544115 | 51,89) | 0,058 | 50,93 (от 25,47 до 76,40) | 0,001 | 25,32 (1,62 до 49,02) | 0,033 | |||||||||
| SGRQ-I 9011 9011 9011 | 125 94612 94612 94612 | 125 | |||||||||||||||||
| T 1 — T 0 | -1.24 ± 8,22 | −1,83 ± 5,77 | −3,90 ± 7,35 | 0,309 | −2,07 (−6,15 до 2,01) | 0,519 | −2,66 (−7,49 до 2,17) | 0,448 − | 911 От 5,03 до 3,85)0,983 | ||||||||||
| T 2 — T 0 | −1,82 ± 7,73 | −3,77 ± 8,16 | −8,13 ± 17,19 | 0,10125 −4-1,00 911 ( ) | 0,490 | −6,31 (от -14,59 до 1,98) | 0.182 | -1,96 (-6,91 до 3,00) | 0,704 | ||||||||||
| T 6 — T 0 | -1,08 ± 12,69 | -2,18 ± 14,03 | 90 -7,03 | 0,288 | −4,85 (от −15,45 до 5,75) | 0,599 | −5,95 (от −16,23 до 4,33) | 0,403 | −1,10 (от −9,44 до 7,24) | 0,983 | |||||||||
| T 1 — T 0 | −2.58 ± 8,51 | −4,74 ± 15,14 | −7,12 ± 10,46 | 0,310 | −2,38 (от −10,48 до 5,72) | 0,850 | −4,54 (от −10,43 до 1,35) | −4,54 (−10,43 до 1,35) | 0,177 — | ,16 От 9,87 до 5,55)0,866 | |||||||||
| T 2 — T 0 | −4,52 ± 13,95 | −11,16 ± 23,75 | −18,42 ± 20,15 | 0,023 −46 до 6,311 ) | 0,477 | −13,90 (от −24,61 до 5,58) | 0.007 | −6,64 (от −18,86 до 5,58) | 0,455 | ||||||||||
| T 6 — T 0 | −3,39 ± 29,56 | −7,68 ± 24,47 | −7,68 ± 24,47 | −915,52 −7,83 (от −21,06 до 5,39) | 0,384 | −12,13 (от −27,30 до 3,04) | 0,152 | −4,30 (от −21,23 до 12,63) | 0,898 | ||||||||||
| SGRQ-I (ударопрочный) | 9115|||||||||||||||||||
| T 1 — T 0 | −5.56 ± 10,98 | −6,82 ± 17,92 | −9,24 ± 13,63 | 0,591 | −2,42 (от −12,30 до 7,47) | 0,907 | −3,68 (от −11,33 до 3,96) | 9116−3,68 (от −11,33 до 3,96) | 9116−0,560 | От 10,58 до 8,05)0,982 | |||||||||
| T 2 — T 0 | −6,20 ± 19,26 | −10,92 ± 24,05 | −21,91 ± 21,47 | 0,025 −15 3,19 до 9113 ) | 0,170 | −15,70 (от −28,30 до −3.11) | 0,010 | −4,72 (от −18,32 до 8,88) | 0,777 | ||||||||||
| T 6 — T 0 | −4,15 ± 27,97 | −9,41 ± 30115 | 25,80,050 | −11,44 (от −28,15 до 5,28) | 0,263 | −16,69 (от −32,54 до −0,85) | 0,036 | −5,27 (от −23,46 до 12,95) | 0,858 | -I1144 (S) Итого)||||||||||
| T 1 — T 0 −3 | ± 8,27 | −5,31 ± 12,96 | −7,61 ± 10,03 | 0,363 | −2,30 (от −9,49 до 4,89) | 0,817 | −3,79 (от −9,47 до 1,89) | 0,284 −1,49 911 От 8,30 до 5,32) | 0,930 | ||||||||||
| T 2 — T 0 | −4,87 ± 14,06 | −9,76 ± 19,26 | −18,42 ± 18,31 −1 | 0,009 −208,66 911 ) | 0.200 | −13,56 (от −23,64 до −3,47) | 0.005 | -4,90 (от -15,43 до 5,64) | 0,586 | ||||||||||
| T 6 — T 0 | -3,35 ± 24,63 | -7,73 ± 22,28 | 4 | -7,73 ± 22,28 | 4 | 0,01 0,01 0,01 0,02 −8,99 (от −21,72 до 3,74) | 0,240 | −13,36 (от −26,93 до 0,20) | 0,055 | −4,37 (от −19,01 до 10,27) | 0,845 | ||||||||
| mMRC12 | 911 946 946|||||||||||||||||||
| T 1 — T 0 | −0.03 ± 0,31 | -0,23 ± 0,43 | -0,22 ± 0,42 | 0,091 | 0,01 (-0,25 до 0,27) | NA | -0,19 (-0,42 до 0,04) | 0,142 | -0,19 ( до 0,04) | 0,131 | |||||||||
| T 2 — T 0 | −0,10 ± 0,40 | −0,42 ± 0,50 | −0,50 ± 0,57 | 911 −46 0,004 9110,40 −0,08 (0,25)0,908 | -0,40 (от -0,71 до -0,10) | 0.005 | -0,32 (-0,60 до 0,04) | 0,020 | |||||||||||
| T 6 — T 0 | -0,06 + 0,68 | -0,45 ± 0,73 | -0,512 9115 9115 0,07 0,07 -0,08 (-0,48 до 0,32) | 0,948 | -0,47 (-0,85 до -0,08) | 0,013 | -0,39 (-0,82 до 0,05) | 0,097 | |||||||||||
| FVC (L) | |||||||||||||||||||
| T 2 — T 0 | −0.02 ± 0,32 | 0,05 ± 0,39 | 0,23 ± 0,36 | 0,020 | 0,18 (от -0,05 до 0,41) | 0,160 | 0,25 (0,04 до 0,46) | 0,016 | 0,06 (от -012,16 до | ) 0,853 | |||||||||
| DL CO (% прогнозируемого) | |||||||||||||||||||
| 2,33 ± 4,35 | 5,10 ± 8,47 | 0,040 | 2,77 (от -1,40 до 6,95) | 0,285 | 4,19 (от -0,37 до 8,76) | 0,080 | 5 от 1,42 до -1,320,646 | ||||||||||||
Как показано на рисунке, общие баллы по симптомам, активности, влиянию и SGRQ-I снизились через 1 и 2 месяца по сравнению с исходным уровнем в трех группах и увеличились через 4 месяца наблюдения по сравнению с концом. вмешательства.Были статистические различия между изменениями по сравнению с исходной активностью, воздействием и общим баллом SGRQ-I через 2 месяца между тремя группами.
Симптомы, активность, влияние и общее количество модуля SGRQ ‐ I с течением времени в ходе исследования. , PD; , упражнение; , контроль. PD, легочный Даоинь; SGRQ ‐ I, Георгиевский респираторный опросник для IPF.
Никаких различий не наблюдалось в отношении симптомов и изменений по сравнению с исходным уровнем через 1 и 6 месяцев для всех доменов SGRQ-I (таблица). Межгрупповая разница в среднем изменении от исходного уровня для PD и контрольной группы для общего балла SGRQ-I составила -13.56 (95% ДИ: от -23,64 до -3,47; P = 0,005) через 2 месяца и -13,36 (95% ДИ: от -26,93 до 0,20; P = 0,055) через 6 месяцев; для удара оно составило -15,70 (95% ДИ: от -28,30 до -3,11; P = 0,010) и -16,69 (95% ДИ: от -32,54 до -0,85; P = 0,036) соответственно; для активности она составила -13,90 (95% ДИ: от -24,61 до 5,58; P = 0,007) через 2 месяца.
Вторичные исходы
Как показано на рисунке, оценка mMRC снизилась через 1 и 2 месяца по сравнению с исходным уровнем в трех группах и увеличилась через 4 месяца наблюдения по сравнению с окончанием вмешательства.Между тремя группами наблюдались статистические различия в изменениях по сравнению с исходным уровнем показателя mMRC через 2 и 6 месяцев. Никаких отличий не наблюдалось для изменений по сравнению с исходным уровнем через 1 месяц (таблица). Межгрупповая разница в среднем изменении от исходного уровня для PD и контроля для оценки mMRC составила -0,40 (95% ДИ: от -0,71 до -0,10; P = 0,005) через 2 месяца и -0,47 (95% ДИ: — От 0,85 до -0,08; P = 0,013) через 6 месяцев; для упражнений и контроля оно составило -0,32 (95% ДИ: от -0,60 до 0.04; P = 0,020) через 2 месяца. Других отличий не наблюдалось.
Как видно на рисунке, произошли изменения в FVC и DL CO (% прогнозируемого). Разница между PD и контрольной группой в среднем изменении от исходного уровня FVC через 2 месяца составила 0,25 (95% ДИ: 0,04–0,46; P = 0,016), и она составила 4,19 для DL CO (прогнозируемый%) ( 95% ДИ: от -0,37 до 8,76; P = 0,080). Не наблюдалось различий для FVC и DL CO (прогнозируемый%) между PD и упражнениями или группами упражнений и контроля (таблица).Парные сравнения для FVC и DL CO (% прогнозируемого) среди трех групп через 2 месяца показали значительную разницу между PD и контрольными группами ( P = 0,046 и 0,006, соответственно; таблица).
Таблица 3
Парные сравнения для FVC и DL CO среди трех групп на втором месяце
| Переменная результата | Группа | Группа | P -значение | 95% ДИ для разницы | ||||
|---|---|---|---|---|---|---|---|---|
| FVC | PD | Упражнение | 0.845 | от −0,256 до 0,473 | ||||
| Контроль | 0,046 | от 0,004 до 0,601 | ||||||
| Упражнение | PD 25 | PD 256 0,86 | Контроль | 0,469 | −0,168 до 0,556 | |||
| DL CO (% прогнозируемого) | PD | Упражнение | 0,631 | −3.923 до 10,071 | ||||
| Контроль | 0,006 | 1,727 до 12,754 | ||||||
| Упражнение | PD | 0,631 | −10,044 | 0,631 | −10,044 | −10.044 0,273 | -2,008 до 10,341 |
Изменения 6MWD, mMRC, FVC и DL CO (прогнозируемый%) по сравнению с исходным уровнем через 1, 2 и 6 месяцев показаны на рисунке.
Безопасность
Всего 10 пациентов (1 в группе PD, 3 в группе упражнений и 6 в контрольной группе) сообщили о нежелательных явлениях (НЯ). Среди них шесть НЯ были отмечены как легкие или умеренные, в том числе два пациента с гипертонией, один пациент с болью в коленях, один пациент с ощущением стеснения в груди, один пациент с простудой и один пациент с острым обострением ИЛФ ( АЕ ‐ IPF). Все пациенты полностью вылечились от НЯ после лечения или прекращения обучения и остались в исследовании.Четыре пациента (один в группе упражнений и три в контрольной группе) испытали тяжелые НЯ. Они погибли из-за AE ‐ IPF.
ОБСУЖДЕНИЕ
Насколько нам известно, наше исследование является первым, в котором изучается влияние программы PR TCM на лечение IPF. Мы обнаружили, что программа PD на 2 месяца по сравнению с группой упражнений и контрольной группой превышала минимальную клинически значимую разницу (MCID) 28 м с диапазоном 10,8–58,5 м. 27 или 25–45 м 28 год 6MWD для пациентов с IPF.Никаких значительных изменений в общей шкале SGRQ-I, mMRC или спирометрии не произошло ни в одной из групп. Мы пришли к выводу, что PD и упражнения не уступают PR, а PD может повысить толерантность к физической нагрузке.
Результаты настоящего исследования демонстрируют, что программа PD была более эффективной, чем контрольная группа или группа упражнений в увеличении 6MWD. Наши результаты согласуются с результатами предыдущих метаанализов. 29 , 30 , 31 год которые изучали эффективность PR в нескольких исследованиях, которые показали, что PR увеличивает толерантность к физической нагрузке у пациентов с IPF.PR — это комплексное вмешательство, предназначенное для улучшения физического и психологического состояния пациента и содействия долгосрочному соблюдению правил поведения, способствующих укреплению здоровья. 32 Основная цель — облегчить симптомы одышки, улучшить дыхательную функцию, улучшить переносимость упражнений, улучшить способность участвовать в общественной и повседневной жизни, снизить частоту госпитализаций и медицинских расходов, а также улучшить общее качество жизни. Руководство IPF ATS / ERS / JRS / ALAT 2015 10 и японские рекомендации по лечению ИЛФ 12 предполагают, что пациентов с IPF в хронической фазе следует лечить PR.
6MWD — полезный показатель функциональной способности, предназначенный для людей с нарушениями как минимум средней степени тяжести. 19 Это полезно для оценки степени тяжести стадии заболевания, предоставляет информацию о терапевтических результатах и прогнозирует заболеваемость и смертность у пациентов с ИЛФ. 28 год , 33 , 34 Общий балл SGRQ является одним из независимых прогностических факторов у пациентов с ИЛФ. 35 год SGRQ-I — полезный измеритель HRQoL у пациентов с IPF. 36 Было обнаружено, что PR улучшает расстояние ходьбы и качество жизни HR у пациентов с ИЛФ.
В нашем исследовании 6MWD увеличились на 60,44 м в группе PD, 32,16 м в группе упражнений и 12,42 м в контрольной группе после 2-месячной программы реабилитации. Значимость наших результатов зависит от MCID измеряемых параметров. Разница между PD и группами упражнений в среднем изменении от исходного уровня для 6MWD через 2 месяца превышала MCID 28 м с диапазоном 10,8–58,5 м. 27 или 25–45 м 28 год 6MWD для пациентов с IPF. Через четыре месяца после окончания вмешательства разница между средним изменением среднего изменения для 6MWD между группами PD и группами упражнений по сравнению с исходным уровнем составила 25.61 м (95% ДИ: от -0,67 до 51,89; P = 0,058). Хотя статистически это не различается, разница в 6MWD между двумя группами составила 25,61 м, что классифицируется как MCID (25–45 м), и в течение 4-месячного периода наблюдения не проводилось никаких тренировок по реабилитации. Межгрупповая разница в среднем изменении от исходного уровня для PD и упражнений для общего балла SGRQ-I через 2 и 6 месяцев превысила MCID (5-8 баллов), 37 но существенных различий между группами не было. Метаанализ семи рандомизированных контролируемых исследований (РКИ) с участием 190 пациентов. 29 пришли к выводу, что PR положительно влияет на 6MWD из 48.60 м (95% ДИ: от 29,03 до 68,18), а общий балл SGRQ-I был ниже (-7,87; 95% ДИ: от -11,44 до -4,30). В другом исследовании наблюдалось среднее снижение на 9,1 общего балла по шкале SGRQ-I для пациентов с IPF после реабилитации в течение 3 месяцев и увеличение на 15,5 в контрольной группе. 38
Одышка — частый симптом, который испытывают пациенты с ИЛФ, и обычно прогрессирует от месяцев до лет. Уменьшение одышки — цель пиара. MMRC отражает одышку у пациентов с IPF, и его можно использовать в качестве простого скринингового инструмента для оценки эффективности.25 Насколько нам известно, MCID по шкале mMRC одышки, FVC и DL CO (прогнозируемый%) у пациентов с IPF не описаны. Разница между группами PD и группами упражнений в среднем изменении оценки одышки по шкале mMRC по сравнению с исходным уровнем через 2 и 6 месяцев статистически не различалась. Эквивалентность оценки одышки по шкале mMRC в группах PD и упражнений в конце вмешательства может быть связана как с увеличением силы четырехглавой мышцы. 39
Спирометрия широко применяется при диагностике, классификации, лечении и мониторинге пациентов с ИЛФ.16 FVC и DL CO (прогнозируемый%) являются типичными параметрами, используемыми в клинических исследованиях. Удивительно, но в нашем исследовании мы обнаружили значительное увеличение FVC и DL CO через 2 месяца только в группе PD. Предыдущие исследования показали, например, снижение DL CO (прогнозируемый%) на 2 для программы тренировок и 4 для контрольной группы через 11 месяцев. 14 Наблюдаемые изменения могут быть связаны с оценкой изменений за относительно короткий период (2 месяца), небольшим размером выборки или исходным состоянием здоровья пациентов.Возможно, что исходный уровень медицинского обслуживания пациентов, участвующих в нашем исследовании, ниже, чем обычно наблюдается в клинических испытаниях IPF. Также возможно, что, поскольку ПД предназначена для улучшения психологических результатов, пациенты в этой группе были более мотивированы, чтобы лучше выполнять эти тесты. Интересно, что существует известная взаимосвязь между депрессией и снижением функции легких. 40 и хотя нам не удалось найти сопоставимых исследований у пациентов с ИЛФ, у пациентов с ХОБЛ, лечившихся от депрессии, были отмечены улучшения в измерениях спирометрии.41 год
6MWD был значительно увеличен в группе PD по сравнению с группой упражнений с величиной, которая превышала MCID. Не было различий в оценке SGRQ-I или оценке одышки по шкале mMRC между группами PD и группами упражнений. Это открытие должно стимулировать PD как замену PR, потому что оно предполагает эффект и низкую стоимость вмешательства программы PD.
PD может работать как вмешательство в IPF за счет следующих механизмов: Пациенты IPF обычно демонстрируют пониженную податливость легких и неэффективные модели дыхания.В этом исследовании программа PD объединила специально разработанные движения конечностей и туловища и контролируемые дыхательные упражнения. Это предназначено для эффективного расширения грудной клетки и растяжения грудных мышц, что может улучшить эффективность дыхания и силу мышц. Менее вероятные, но осуществимые варианты заключаются в том, что ПД увеличивает эластичность плевры, улучшает вентиляционную способность легких и увеличивает объем вентиляции легких.
Наше исследование имеет некоторые ограничения. Во-первых, продолжительность была короткой.Мы наблюдали некоторые тенденции в наших данных, которые могли значительно отличаться, если бы продолжительность исследования была больше. Во-вторых, несмотря на единообразное обучение участников группы ПД, было отмечено, что каждый пациент имеет разный уровень подготовки, что может повлиять на индивидуальную эффективность ПД. В эту исследуемую популяцию не принимали ни один из противофиброзных препаратов пирфенидон и нинтеданиб. Следовательно, комбинированные эффекты антифиброза и PD необходимо оценить в других будущих исследованиях.
В заключение, клинически значимая разница для 6MWD наблюдалась через 2 месяца PD. Результаты исследования показывают, что программа PD является подходящей альтернативой PR при лечении IPF.
Заявление о раскрытии информации
Управление клиническими данными осуществляла компания Jiangsu Famous Medical Technology Co., Ltd, Нанкин, Китай. Компания не участвовала в разработке исследования, интерпретации данных и написании этой рукописи. Эти три клинических центра включали Третью дочернюю больницу Хэнаньского университета китайской медицины, больницу китайской медицины Ручжоу и дочернюю больницу Чанчуньского университета китайской медицины.
Вклад авторов
Концептуализация: H.Z., J.L. Обработка данных: M.Z., H.Z., F.L., Z.Y., C.Y., B.O., J.L. Формальный анализ: H.Z., B.O. Финансирование: М.З. Руководство проекта: М.З. Исследование: F.L., Z.Y., C.Y. Письмо — черновик: М.З., Х.З. Написание — просмотр и редактирование: B.O., J.L.
Благодарности
Мы благодарим всех коллег, научных сотрудников, респираторных врачей и тех, кто поддержал это исследование. Это исследование было поддержано Программой исследовательского проекта по развитию практики национальных клинических исследовательских баз традиционной китайской медицины (грант No.JDZX2015158), докторский фонд Хэнаньского колледжа традиционной китайской медицины, Китай (грант № BSJJ2015-27) и Специальной программы национальных клинических исследовательских баз традиционной китайской медицины администрации провинции Хэнань, Китай (грант № 2018JDZX007). Источники финансирования не играли никакой роли ни в разработке и проведении исследования, ни в написании рукописи для публикации.
Банкноты
Чжоу М., Чжан Х., Ли Ф. и др. Легочный Даоинь как программа реабилитации пациентов традиционной китайской медицины для пациентов с ИЛФ: рандомизированное контролируемое исследование.Респирология. 2021; 26: 360–369. 10.1111 / соотв.13972 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Регистрация клинического исследования: ChiCTR ‐ IOR ‐ 17011187 на сайте www.chictr.org.cn
(Заместитель редактора: Мелисса Бентон; старший редактор: Крис Грейндж)
Заявление о доступности данных
Индивидуальные данные участников, которые лежат в основе результатов, представленных в этой статье, и протокол исследования могут быть доступны после деидентификации (текст, таблицы, рисунки и приложения) от 9 до 36 месяцев после публикации статьи для метаанализа данных отдельных участников.Предложения исследователя об использовании данных должны быть одобрены независимым комитетом по анализу, назначенным для этой цели с подписанным соглашением о доступе к данным.
ССЫЛКИ
1. Naccache JM, Montil M, Cadranel J, Cachanado M, Cottin V, Crestani B, Valeyre D, Wallaert B, Simon T., Nunes H. Протокол исследования: изучение эффективности циклофосфамида, добавленного к кортикостероидам, для лечения острого обострения идиопатического фиброза легких; рандомизированное двойное слепое плацебо-контролируемое многоцентровое исследование III фазы (EXAFIP).BMC Pulm. Med. 2019; 19: 75. [Бесплатная статья PMC] [PubMed] [Google Scholar] 2. Баддини ‐ Мартинес Дж., Перейра, Калифорния. Сколько пациентов с идиопатическим фиброзом легких в Бразилии? J. Bras. Пневмол. 2015; 41: 560–1. [Бесплатная статья PMC] [PubMed] [Google Scholar] 3. Harari S, Madotto F, Caminati A, Conti S, Cesana G. Эпидемиология идиопатического легочного фиброза в Северной Италии. PLoS One 2016; 11: e0147072. [Бесплатная статья PMC] [PubMed] [Google Scholar] 4. Ли ХЭ, Мён ДжейПи, Ким Х.Р., Ри СК, Юн ХК, Ку ДжВ.Заболеваемость и распространенность идиопатической интерстициальной пневмонии и идиопатического фиброза легких в Корее. Int. J. Tuberc. Lung Dis. 2016; 20: 978–84. [PubMed] [Google Scholar] 5. Нацуизака М., Чиба Х., Куронума К., Оцука М., Кудо К., Мори М., Бандо М., Сугияма Й., Такахаши Х. Эпидемиологическое обследование японских пациентов с идиопатическим фиброзом легких и исследование этнических различий. Являюсь. J. Respir. Крит. Care Med. 2014; 190: 773–9. [PubMed] [Google Scholar] 6. Рагху Г., Чен С.Ю., Хоу К., Йе В.С., Коллард Х.Р.Заболеваемость и распространенность идиопатического фиброза легких у взрослых в США в возрасте 18–64 лет. Евро. Респир. Дж. 2016; 48: 179–86. [PubMed] [Google Scholar] 7. Мартинес Ф.Дж., Коллард Х.Р., Пардо А., Рагху Дж., Ричелди Л., Селман М., Свигрис Дж. Дж., Танигучи Х., Уэллс, Австралия. Идиопатический фиброз легких. Nat. Преподобный Дис. Грунтовки. 2017; 3: 17074. [PubMed] [Google Scholar] 8. Рагху Г., Чен С.Ю., Йе В.С., Марони Б., Ли Кью, Ли Ю.С., Коллард HR. Идиопатический фиброз легких у получателей программы Medicare в США в возрасте 65 лет и старше: заболеваемость, распространенность и выживаемость, 2001-11 гг.Ланцет Респир. Med. 2014; 2: 566–72. [PubMed] [Google Scholar] 9. Ледерер DJ, Мартинес FJ. Идиопатический фиброз легких. N. Engl. J. Med. 2018; 378: 1811–23. [PubMed] [Google Scholar] 10. Рагху Дж., Рохверг Б., Чжан И., Гарсия, Калифорния, Адзума А., Бер Дж., Брозек Дж. Л., Коллард Х. Р., Каннингем В., Хомма С. et al., .; Американское торакальное общество; Европейское респираторное общество; Японское респираторное общество; Латиноамериканская торакальная ассоциация. Официальное руководство по клинической практике ATS / ERS / JRS / ALAT: лечение идиопатического фиброза легких: обновление руководства по клинической практике 2011 года.Являюсь. J. Respir. Крит. Care Med. 2015; 192: e3–19. [PubMed] [Google Scholar] 11. Рагху Дж., Реми-Жардин М., Майерс Дж. Л., Ричелди Л., Райерсон С. Дж., Ледерер Д. Д., Бер Дж., Коттин В., Данофф С. К., Морелл Ф. и др. . Диагностика идиопатического фиброза легких. Официальное руководство по клинической практике ATS / ERS / JRS / ALAT. Являюсь. J. Respir. Крит. Care Med. 2018; 198: e44–68. [PubMed] [Google Scholar] 12. Хомма С., Бандо М., Адзума А., Сакамото С., Сугино К., Исии Ю., Изуми С., Инасе Н., Иноуэ Ю., Эбина М. и др. .Японские рекомендации по лечению идиопатического фиброза легких. Респир. Расследование. 2018; 56: 268–91. [PubMed] [Google Scholar] 13. Феррейра А., Гарви С., Коннорс Г.Л., Хиллинг Л., Риглер Дж., Фаррелл С., Кайу К., Шариат С., Коллард Х.Р. Легочная реабилитация при интерстициальной болезни легких; преимущества и предикторы ответа. Грудь 2009; 135: 442–7. [PubMed] [Google Scholar] 14. Vainshelboim B, Oliveira J, Fox BD, Soreck Y, Fruchter O, Kramer MR. Долгосрочное влияние 12-недельной программы тренировок на клинические исходы при идиопатическом фиброзе легких.Легкое 2015; 193: 345–54. [PubMed] [Google Scholar] 15. Доуман Л.М., Макдональд С.Ф., Хилл С.Дж., Ли А.Л., Баркер К., Бут С., Гласпоул I, Го Н.С.Л., Сауткотт А.М., Бердж А.Т. и др. . Доказательства преимуществ физических упражнений при интерстициальном заболевании легких: рандомизированное контролируемое исследование. Грудная клетка 2017; 72: 610–9. [PubMed] [Google Scholar] 16. Ю XQ, Ли JS, Ли SY, Се Y, Ван М. Х., Чжан Х. Л., Ван Х. Ф., Ван Ц. В. Функциональные и психосоциальные эффекты легочного даоиня на пациентов с ХОБЛ в Китае: протокол многоцентрового рандомизированного контролируемого исследования.J. Integr. Med. 2013; 11: 140–6. [PubMed] [Google Scholar] 17. Zhang HL, Li JS, Yu XQ, Li SY, Halmurat U, Xie Y, Wang YF, Li FS, Wang MH. Оценка переносимости активности, результатов, сообщаемых пациентами, и удовлетворенности эффективностью легочного даоиня у пациентов с хронической обструктивной болезнью легких. Int. J. Chron. Препятствовать. Легочный. Дис. 2017; 12: 2333–42. [Бесплатная статья PMC] [PubMed] [Google Scholar] 18. Джексон Р.М., Гомес-Марин О.В., Рамос К.Ф., Соль К.М., Коэн М.И., Гаунаурд И.А., Кахалин Л.П., Карденас Д.Д.Ограничение физических упражнений у пациентов с IPF: рандомизированное исследование легочной реабилитации. Легкое 2014; 192: 367–76. [PubMed] [Google Scholar] 19. Комитет ATS по профессиональным стандартам для лабораторий клинических исследований функции легких . Заявление ATS: рекомендации для теста с шестиминутной ходьбой. Являюсь. J. Respir. Крит. Care Med. 2002; 166: 111–7. [PubMed] [Google Scholar] 20. Prior TS, Hoyer N, Shaker SB, Davidsen JR, Yorke J, Hilberg O, Bendstrup E. Проверка версии респираторного опросника Святого Георгия, специфичной для IPF.Респир. Res. 2019; 20: 199. [Бесплатная статья PMC] [PubMed] [Google Scholar] 21. Glaspole IN, Chapman SA, Cooper WA, Ellis SJ, Goh NS, Hopkins PM, Macansh S, Mahar A, Moodley YP, Paul E и др. . Связанное со здоровьем качество жизни при идиопатическом фиброзе легких: данные австралийского реестра IPF. Респирология 2017; 22: 950–6. [PubMed] [Google Scholar] 22. Натан С.Д., Шлобин О.А., Вейр Н., Ахмад С., Калджоб Джобс, Баттл Э., Шеридан М.Дж., дю Буа Р.М. Отдаленное течение и прогноз идиопатического фиброза легких в новом тысячелетии.Грудь 2011; 140: 221–9. [PubMed] [Google Scholar] 23. Малер Д.А., Уэллс СК. Оценка клинических методов оценки одышки. Грудь 1988; 93: 580–6. [PubMed] [Google Scholar] 24. Bestall JC, Paul EA, Garrod R, Garnham R, Jones PW, Wedzicha JA. Полезность шкалы одышки Медицинского исследовательского совета (MRC) в качестве показателя инвалидности у пациентов с хронической обструктивной болезнью легких. Грудная клетка 1999; 54: 581–6. [Бесплатная статья PMC] [PubMed] [Google Scholar] 25. Rajala K, Lehto JT, Sutinen E, Kautiainen H, Myllärniemi M, Saarto T.Шкала одышки mMRC указывает на ухудшение качества жизни и усиление боли у пациентов с идиопатическим фиброзом легких. ERJ Open Res. 2017; 3: 00084‐2017. [Бесплатная статья PMC] [PubMed] [Google Scholar] 26. Takeda T, Takeuchi M, Saitoh M, Takeda S. Улучшение результатов, сообщаемых пациентами, и увеличение жизненной емкости легких во время лечения идиопатического легочного фиброза нинтеданибом. Tohoku J. Exp. Med. 2018; 245: 107–14. [PubMed] [Google Scholar] 27. Свигрис Дж. Дж., Вамбольдт Ф. С., Бер Дж., Дю Буа Р. М., Кинг Т. Э., Рагху Дж., Браун К. К..6-минутная прогулка при идиопатическом фиброзе легких: продольные изменения и минимально важная разница. Грудная клетка 2010; 65: 173–7. [Бесплатная статья PMC] [PubMed] [Google Scholar] 28. дю Буа Р.М., Вейкер Д., Альбера С., Брэдфорд В.З., Костабель Ю., Карташов А., Ланкастер Л., Нобл П. В., Сан С. А., Шварцберг Дж. и др. . Тест с шестиминутной ходьбой при идиопатическом фиброзе легких: валидация теста и минимальная клинически значимая разница. Являюсь. J. Respir. Крит. Care Med. 2011; 183: 1231–7. [PubMed] [Google Scholar] 29.Yu X, Li X, Wang L, Liu R, Xie Y, Li S, Li J. Легочная реабилитация для толерантности к физической нагрузке и качества жизни у пациентов с IPF: систематический обзор и метаанализ. Биомед. Res. Int. 2019; 2019: 8498603. [Бесплатная статья PMC] [PubMed] [Google Scholar] 30. Гомес ‐ Нето М., Сильва С.М., Эсекьель Д., Консейсао К.С., Сакетто М., Мачадо А.С. Влияние легочной реабилитации на переносимость физической нагрузки и качество жизни пациентов с идиопатическим фиброзом легких: систематический обзор и метаанализ. J. Cardiopulm.Rehabil. Пред. 2018; 38: 273–8. [PubMed] [Google Scholar] 31. Cheng L, Tan B, Yin Y, Wang S, Jia L, Warner G, Jia G, Jiang W. Краткосрочные и долгосрочные эффекты легочной реабилитации при идиопатическом фиброзе легких: систематический обзор и метаанализ. Clin. Rehabil. 2018; 32: 1299–307. [PubMed] [Google Scholar] 32. Спруит М.А., Сингх С.Дж., Гарви К., ЗуУоллак Р., Ники Л., Рочестер К., Хилл К., Голландия А.Е., Ларо СК, Мэн В.Д. и др. . Официальное заявление Американского торакального общества / Европейского респираторного общества: ключевые концепции и достижения в легочной реабилитации.Являюсь. J. Respir. Крит. Care Med. 2013; 188: e13–64. [PubMed] [Google Scholar] 33. Lancaster LH. Полезность теста с шестиминутной ходьбой у пациентов с идиопатическим фиброзом легких. Многопрофильный. Респир. Med. 2018; 13:45 [Бесплатная статья PMC] [PubMed] [Google Scholar] 34. Хереси Г.А., Двейк Р.А.. Сильные и слабые стороны теста с шестиминутной ходьбой: исследование модельных биомаркеров идиопатического легочного фиброза. Являюсь. J. Respir. Крит. Care Med. 2011; 183: 1122–4. [PubMed] [Google Scholar] 35. Фурукава Т., Танигучи Х., Андо М., Кондо Й., Катаока К., Нишияма О, Джоко Т, Фукуока Дж., Сакамото К., Хасэгава Ю.Георгиевский респираторный опросник как прогностический фактор при ИЛФ. Респир. Res. 2017; 18: 18. [Бесплатная статья PMC] [PubMed] [Google Scholar] 36. Йорк Дж., Джонс П. В., Свигрис Дж. Дж. Разработка и проверка достоверности версии респираторного опросника Святого Георгия для IPF. Грудная клетка 2010; 65: 921–6. [Бесплатная статья PMC] [PubMed] [Google Scholar] 37. Свигрис Дж. Дж., Эссер Д., Коносенти К. С., Браун К. К.. Психометрические свойства респираторного опросника Святого Георгия (SGRQ) у пациентов с идиопатическим фиброзом легких: обзор литературы.Качество здоровья. Результаты жизни 2014; 12: 124. [Бесплатная статья PMC] [PubMed] [Google Scholar] 38. Гаунаурд И.А., Гомес-Марин О.В., Рамос К.Ф., Сол С.М., Коэн М.И., Кахалин Л.П., Карденас Д.Д., Джексон Р.М. Улучшение физической активности и качества жизни пациентов с идиопатическим фиброзом легких, завершающих программу легочной реабилитации. Респир. Забота 2014; 59: 1872–9. [PubMed] [Google Scholar] 39. Келли JL, Элкин SL, Fluxman J, Polkey MI, Soljak MA, Hopkinson NS. Одышка и слабость скелетных мышц у пациентов, проходящих скрининг на здоровье легких в первичной медико-санитарной помощи.ХОБЛ 2013; 10: 40–54. [PubMed] [Google Scholar] 40. Ochs ‐ Balcom HM, Lainhart W., Mnatsakanova A, Charles LE, Violanti JM, Andrew ME, Freudenheim JF, Muti P, Trevisan M, Burchfiel CM и др. . Связь депрессивных симптомов и легочной функции у здоровых взрослых. Психосом. Med. 2013; 75: 737–43. [Бесплатная статья PMC] [PubMed] [Google Scholar] 41. Момтаз ОМ, Рабей С.М., Тауфик Н.Р., Хасан А.А. Влияние лечения депрессии и тревоги на физиологическое состояние пациентов с тяжелой формой ХОБЛ.Египет. J. Chest Dis. Tuberc. 2015; 64: 29–34. [Google Scholar]Открытие механизма двойной протеазы, который способствует восстановлению контрольных точек повреждений ДНК
Abstract
Ответ на повреждение ДНК — это сигнальный путь, обнаруженный во всей биологии. У многих бактерий контрольная точка повреждения ДНК обеспечивается за счет индукции экспрессии небольшого мембраносвязанного ингибитора, который задерживает деление клеток, обеспечивая время для восстановления поврежденных хромосом. Как клетки способствуют восстановлению контрольной точки после обнаружения успешного восстановления, неизвестно.Используя высокопроизводительный прямой генетический скрининг, мы идентифицировали две неродственные протеазы, YlbL и CtpA, которые способствуют восстановлению контрольной точки повреждения ДНК в Bacillus subtilis . Делеция обеих протеаз приводит к накоплению белка контрольной точки YneA. Мы показываем, что чувствительность к повреждению ДНК и увеличенное удлинение клеток у мутантов протеазы зависит от yneA . Кроме того, экспрессия YneA в мутантах протеазы была достаточной для подавления пролиферации клеток. Наконец, мы показываем, что обе протеазы взаимодействуют с YneA и что одна из двух протеаз, CtpA, непосредственно расщепляет YneA in vitro .С этими результатами мы сообщаем о механизме восстановления контрольной точки повреждения ДНК у бактерий, которые используют мембраносвязанные ингибиторы деления клеток.
Сведения об авторе
Прокариоты и эукариоты координируют деление клеток с соблюдением целостности генома, используя контрольные точки повреждения ДНК. Многие бактерии экспрессируют небольшой мембранно-связывающий белок, замедляющий деление клеток, когда встречаются препятствия для репликации ДНК. Ингибиторы клеточного деления этого класса были идентифицированы у нескольких видов бактерий, но механизм, используемый для ослабления ингибирования, остается неизвестным.Используя прямую генетику, мы идентифицировали два неизученных гена, кодирующих протеазы YlbL и CtpA, удаление которых приводит к чувствительности к лекарствам, которые непосредственно повреждают ДНК. Мы показываем, что чувствительность к повреждению ДНК у мутантов протеазы является результатом накопления ингибитора деления клетки. Кроме того, мы показываем, что YlbL и CtpA ответственны за деградацию ингибитора клеточного деления, позволяя возобновить клеточное деление. Важно отметить, что эти две протеазы не являются гомологами, что является ярким примером того, как бактерия использует негомологичные ферменты для разложения одного субстрата.Наше исследование раскрывает ранее неизвестный механизм, используемый для удаления ингибитора деления клеток, а также освещает потенциальную стратегию, которую бактерии могут использовать для регулирования сигнальных путей. Использование множества несвязанных белков для выполнения одной функции может представлять собой стратегию, применяемую во всех биологических системах.
Образец цитирования: Burby PE, Simmons ZW, Schroeder JW, Simmons LA (2018) Открытие механизма двойной протеазы, который способствует восстановлению контрольной точки повреждения ДНК.PLoS Genet 14 (7): e1007512. https://doi.org/10.1371/journal.pgen.1007512
Редактор: Иван Матич, Парижский университет Декарта, INSERM U1001, ФРАНЦИЯ
Поступила: 8 мая 2018 г .; Одобрена: 23 июня 2018 г .; Опубликовано: 6 июля 2018 г.
Авторские права: © 2018 Burby et al. Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Доступность данных: Все соответствующие данные находятся в документе и его файлах вспомогательной информации с данными секвенирования, доступными с использованием инвентарного номера GSE109366. https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE109366
Финансирование: Эта работа была поддержана Национальными институтами здравоохранения (https: //www.nigms.nih .gov / Pages / default.aspx) предоставить LAS R01 GM107312. JWS частично поддерживался Национальными институтами здравоохранения (https://www.nigms.nih.gov/Pages/default.aspx) T32 GM007544 и средства доцента из Колледжа литературы, науки и искусств Мичиганского университета (https://lsa.umich.edu) в LAS. PEB был поддержан стипендией для докторантуры от Национального научного фонда (отдел последипломного образования 1256260). Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили, что никаких конкурирующих интересов не существует.
Введение
Ответ на повреждение ДНК (DDR, SOS-ответ у бактерий) является важным путем для поддержания целостности генома во всех сферах жизни. Неправильная регуляция DDR у людей может приводить к различным болезненным состояниям [1, 2], а у бактерий было обнаружено, что SOS-ответ важен для выживания при многих стрессовых факторах [3-5]. Ответ на повреждение ДНК у всех организмов приводит к трем основным результатам: ответ транскрипции, который может варьироваться в зависимости от типа повреждения ДНК, репарация ДНК и активация контрольной точки повреждения ДНК [6–9].У эукариот контрольные точки G1 / S и G2 / M устанавливаются киназами контрольных точек, которые передают сигнал о повреждении ДНК посредством инактивации фосфатазы Cdc25 [7]. Ингибирование Cdc25, зависимое от киназы контрольных точек, ведет к накоплению фосфорилированных циклинзависимых киназ, что предотвращает прогрессирование клеточного цикла [7]. У бактерий контрольная точка SOS-зависимого повреждения ДНК зависит от экспрессии ингибитора клеточного деления, хотя тип ингибитора варьируется в зависимости от вида бактерий.
В Escherichia coli , SOS-зависимая контрольная точка повреждения ДНК является наиболее изученной бактериальной контрольной точкой [6].При активации SOS-ответа экспрессируется ингибитор деления цитоплазматических клеток SulA [10]. Накопление SulA приводит к блокированию образования перегородки, препятствуя сборке цитокинетического кольца с помощью FtsZ, гомолога эукариотического тубулина [11, 12]. SulA напрямую связывается с FtsZ [13] и ингибирует полимеризацию FtsZ [14, 15]. Восстановление из контрольной точки, индуцированной SulA, происходит посредством протеолиза SulA. Lon является первичной протеазой, ответственной за очищение SulA [16-18], хотя было обнаружено, что ClpYQ (HslUV) вносит вклад в деградацию SulA в отсутствие Lon [19-21].Таким образом, механизмы активации контрольной точки повреждения ДНК цитоплазматическим белком SulA и последующего восстановления хорошо изучены в E . coli . Однако зона проверки, зависящая от SulA, ограничена E . coli и подмножество близкородственных бактерий. Становится все более очевидным, что большинство других бактерий используют контрольную точку повреждения ДНК с совершенно другим механизмом принуждения и восстановления.
Было показано, что эволюционно широкая группа бактериальных организмов использует заметно отличающийся механизм контрольной точки повреждения ДНК [22-25].У этих грамположительных и грамотрицательных организмов экспрессируется небольшой белок с трансмембранным доменом, который ингибирует деление клеток без нацеливания на FtsZ. Одним из примеров является грамотрицательная бактерия Caulobacter crescentus , где белки SidA и DidA связываются с основными мембраносвязанными компонентами дивизом, FtsW / N, которые вносят вклад в ремоделирование пептидогликана [22, 26]. Другой пример — грамположительная бактерия Bacillus subtilis , у которой ингибитор SOS-зависимого деления клеток — YneA [23].YneA содержит N-концевой трансмембранный домен, большая часть белка которого находится во внеклеточном пространстве [27]. После активации SOS LexA-зависимая репрессия yneA снимается и экспрессируется yneA [23]. Повышенная экспрессия yneA приводит к удлинению клеток, хотя образование кольца FtsZ все еще происходит [27], предполагая, что YneA ингибирует деление клеток посредством механизма, отличного от механизма SulA. Дальнейшие исследования показали, что сверхэкспрессированный YneA высвобождается в среду, и что полноразмерный YneA, вероятно, является активной формой белка [27].Механизм (ы), ответственный за инактивацию YneA, неизвестен. Таким образом, хотя использование небольшого, связанного с мембраной ингибитора деления клеток широко распространено среди бактерий, во всех изученных случаях механизм восстановления контрольных точек остается неизвестным [22–26].
Мы сообщаем о серии прямых генетических скринингов трех различных классов агентов, повреждающих ДНК, с использованием транспозонного мутагенеза с последующим глубоким секвенированием (Tn-seq). Наш скрининг выявил две протеазы, YlbL и CtpA, которые важны для роста при наличии повреждений ДНК.Механистические исследования показывают, что YlbL и CtpA имеют перекрывающиеся функции, и в отсутствие этих двух протеаз растяжение клеток, зависящее от повреждения ДНК, увеличивается, а восстановление контрольных точек замедляется. Протеомный анализ выявил накопление YneA в мутанте с двойной протеазой. Мы также обнаружили, что чувствительность к повреждению ДНК мутантов протеазы зависит исключительно от yneA . Кроме того, мы показываем, что обе протеазы взаимодействуют с полноразмерным YneA в бактериальном двухгибридном анализе и что CtpA способна переваривать YneA в очищенной системе.С этими результатами мы представляем модель восстановления контрольной точки повреждения ДНК для бактерий, которые используют более распространенный механизм с использованием небольшого мембраносвязанного ингибитора деления клеток.
Результаты
Обоснование и анализ перспективного генетического скрининга
Чтобы лучше понять реакцию на повреждение ДНК у бактерий, мы выполнили три прямых генетических скрининга с использованием B . subtilis . Мы создали библиотеку вставок транспозонов, состоящую из более чем 120000 различных вставок (таблица S1).Охват каждого транспозона-мутанта в библиотеке был нанесен на график в зависимости от координат генома, что показало, что распределение вставок было приблизительно равномерным по хромосоме в популяции мутантов (рис. 1А). Было обнаружено два небольших исключения, когда охват снизился. Снижение охвата соответствует регионам, в которых сгруппировано много важных генов (рис. 1A, стрелки). С целью идентификации мутантов, важных для ответа на повреждение ДНК, мы выращивали параллельные культуры либо контрольной, либо обработки повреждений ДНК в течение трех периодов роста, моделируя наш экспериментальный план после предыдущего отчета [28; Рис. 1B].Митомицин C (MMC), метилметансульфонат (MMS) и флеомицин (Phleo) были выбраны для скрининга, потому что эти агенты представляют три разных класса антибиотиков, которые непосредственно повреждают ДНК. MMC вызывает меж- и внутрицепочечные сшивки и более крупные аддукты [29, 30], MMS вызывает более мелкие аддукты, состоящие из метилирования ДНК [31], а Phleo приводит к одно- и двухцепочечным разрывам [32, 33]. В результате мы пришли к выводу, что объединенные данные предоставят набор генов, которые обычно важны для ответа на повреждение ДНК.
Рис. 1. Планирование экспериментов по генетическому скринингу и анализ данных.
(A) График логарифма 10 инсерционного покрытия по оси y и геномного положения в нуклеотидах по оси x. (B) Экспериментальный план для экспериментов Tn-seq. Библиотеку транспозонов использовали для инокуляции заквасок, чтобы позволить культурам достичь экспоненциальной фазы. Культуры разделяли на контрольную и обрабатываемую и выращивали в течение трех периодов роста. (C) Уравнения, используемые для расчета относительной приспособленности (W, приспособленность; G, поколения, N, количество клеток в начале (N 0 ) или конце (N f ) периода роста; F, частота вставки в начале (F 0 ) или в конце (F f ) периода роста; n — количество вставок, использованных для расчета среднего). (D) Средняя относительная пригодность гена представлена в виде гистограммы для генов, указанных для всех трех экспериментов с Tn-seq, полосы ошибок представляют 95% доверительный интервал. (E) Диаграмма Венна, изображающая перекрытия 200 генов с наименьшей приспособленностью и скорректированным значением p менее 0,01 для всех трех экспериментов Tn-seq во втором периоде роста.
https://doi.org/10.1371/journal.pgen.1007512.g001
После секвенирования мы провели анализ контроля качества.Во-первых, учитывая, что данные секвенирования являются данными подсчета, распределение покрытия должно быть логарифмически нормальным [34]. Действительно, распределение каждой реплики для образцов исходной библиотеки и заквасочной культуры примерно логарифмически нормальное (S1A фиг.). Мы также обнаружили, что распределения для оставшихся временных точек объединенных реплик соответствовали приблизительному логарифмически-нормальному распределению (S1B, рис.). Данные секвенирования и данные количества жизнеспособных клеток (таблица S1) были использованы для расчета пригодности каждого инсерционного мутанта в каждом состоянии [Рис. 1C; 35, 36].Относительную пригодность каждой вставки рассчитывали путем взятия отношения лечения к контролю (рис. 1С), тем самым выделяя эффекты пригодности обработок. Относительную приспособленность каждого гена определяли путем усреднения относительной приспособленности, рассчитанной для каждой вставки в ген (рис. 1С). Чтобы проверить, подходит ли t-критерий для определения относительной приспособленности, значительно отклоняющейся от единицы, мы построили график распределения относительной приспособленности к вставке. Все распределения были нормальными со средним значением, близким к единице (S1C, рис.).Мы определили относительную приспособленность для каждого гена с достаточным количеством данных (см. Дополнительные методы) и сообщаем значения относительной приспособленности и скорректированные p-значения [37] в таблице S2.
Tn-seq идентифицировал гены, участвующие в репарации ДНК, и гены с неизвестной функцией
Первоначальная проверка показала, что несколько генов, которые, как известно, участвуют в репарации ДНК ( recA , ruvAB , recN и recOR [38]), снизили относительную приспособленность в период роста в одном из всех экспериментов (таблица S2) .Более тщательный анализ трех генов recN , addA и polA , которые находятся в верхней части списков во всех вариантах лечения, показал, что относительная приспособленность меньше единицы в большинстве случаев, хотя в эксперименте Phleo похоже, что культуры адаптировались к обработке к третьему периоду роста (рис. 1D). Для сравнения мы также построили график относительной приспособленности thrC , гена, участвующего в биосинтезе треонина, и обнаружили, что относительная приспособленность составляет примерно единицу во всех исследованных условиях (рис. 1D).Важно отметить, что вставка в uvrA , компонента механизма эксцизионной репарации нуклеотидов [38, 39], который помогает восстанавливать аддукты MMC, но не MMS или повреждения, связанные с флео [40, 41], снижает относительную приспособленность во 2 и 3 периодах роста с MMC. , но не привело к значительному снижению относительной приспособленности в MMS или Phleo (рис. 1D и таблица S2). Взятые вместе, эти результаты подтверждают подход, демонстрируя, что мы смогли идентифицировать гены, которые, как известно, участвуют в репарации ДНК.
Мы также поинтересовались, содержат ли наши результаты ложные срабатывания.Чтобы проверить это, мы решили экспериментально проверить гены с сорока наименьшими значениями относительной приспособленности из периода роста два в эксперименте MMC. Мы обнаружили, что восемь из сорока делеций генов не были чувствительны к MMC в тесте на точечный титр (таблица 1). Несколько генов, которые дали ложноположительные результаты, расположены в геноме рядом с генами с подтвержденными фенотипами, что позволяет предположить, что полярные эффекты объясняют некоторые из ложноположительных результатов (таблица 1; подробный анализ см. В дополнительных результатах). Чтобы идентифицировать гены, которые обычно необходимы как часть ответа на повреждение ДНК, мы исследовали 200 генов из второго периода роста с наименьшей относительной приспособленностью и скорректированным значением p меньше 0.01 из всех трех экспериментов (таблица S3). Мы обнаружили, что 21 ген перекрывается во всех трех экспериментах (рис. 1E), некоторые из которых, как известно, участвуют в репарации ДНК ( recN , addB , polA , radA ), в то время как несколько генов не известны. функция ( e . g ., ylbL и ctpA ) (таблица S3).
Таблица 1. Tn-seq дает много ложноположительных результатов.
Перечислены сорок генов с наименьшей относительной приспособленностью во втором периоде роста в эксперименте MMC Tn-seq.Каждый ген был удален, и делеционные мутанты были протестированы на чувствительность к MMC с использованием анализа титра пятна и диапазона концентраций MMC. Гены, помеченные как нечувствительные, не имели разницы в росте относительно штамма WT на среде, содержащей MMC, за исключением ylbK , что приводило к полярному эффекту на ylbL (см. S3 фиг.).
https://doi.org/10.1371/journal.pgen.1007512.t001
YlbL и CtpA требуют предполагаемых каталитических остатков для функции
Среди генов, важных для роста при наличии повреждений ДНК, мы сосредоточили внимание на двух предполагаемых протеазах YlbL и CtpA.Предполагается, что YlbL имеет три домена: трансмембранный домен, домен Lon-протеазы и домен PDZ (рис. 2A). Предполагается, что CtpA имеет четыре домена: трансмембранный домен, домен пептидазы S41, домен PDZ и C-концевой домен связывания пептидогликана (PG) (рис. 2A). Во всех трех экспериментах с Tn-seq относительная пригодность вставок либо в ylbL , либо в ctpA была значительно меньше, чем одна во второй и третий периоды роста (рис. 2B), что позволяет предположить, что отсутствие любой из протеаз приводит к чувствительности к ДНК. повреждать.Напротив, контрольный ген amyE , который участвует в утилизации крахмала, имел относительную пригодность примерно к единице во всех исследованных условиях (фиг. 2B). Чтобы проверить результаты Tn-seq, мы сконструировали чистые делеции ylbL и ctpA и обнаружили, что оба мутанта чувствительны к повреждению ДНК в тесте на точечный титр (рис. 2С). Каждый фенотип также дополнялся эктопической экспрессией каждой протеазы на соответствующем мутантном фоне (рис. 2С). Чтобы идентифицировать предполагаемые каталитические остатки, мы выровняли протеазный домен YlbL с LonA и LonB из B . subtilis и Lon из E . coli . Выравнивание последовательностей показало, что YlbL содержит предполагаемую каталитическую диаду, состоящую из серина (S234) и лизина (K279) (S2A фиг.). Аналогичным образом мы выровняли CtpA с его гомологами CtpB из B . subtilis и Prc из E . coli , который идентифицировал предполагаемую каталитическую триаду, состоящую из серина (S297), лизина (K322) и глутамина (Q326) (S2B фиг.). Чтобы проверить, необходимы ли эти предполагаемые каталитические остатки для функционирования, мы попытались дополнить фенотип чувствительности к повреждению ДНК посредством эктопической экспрессии мутантов серина и лизина.Как сериновые, так и лизиновые мутанты YlbL и CtpA не смогли дополнить фенотипы делеции (рис. 2С). Варианты и протеазы дикого типа эктопически экспрессировались до одного и того же уровня in vivo (фиг. 2D), что позволяет предположить, что отсутствие комплементации не является следствием нестабильности, вызванной аминокислотными изменениями. На основании этих результатов мы заключаем, что протеазная активность необходима для того, чтобы YlbL и CtpA функционировали в ответ на повреждение ДНК.
Рис. 2. Для функционирования YlbL и CtpA требуются предполагаемые каталитические остатки.
(A) Схема белков YlbL и CtpA, изображающая организацию домена. (B) Средняя относительная пригодность гена представлена в виде гистограммы для amyE , ylbL и ctpA для всех трех экспериментов Tn-seq, столбики ошибок представляют 95% доверительный интервал. (C) Спотовые анализы титра с использованием указанных генотипов и высевают на агаризованную среду LB, содержащую указанные лекарственные средства. (D) Вестерн-блот-анализ клеточных лизатов указанных генотипов, выращенных с ксилозой или без нее, с использованием антисыворотки против YlbL (верхняя панель) и DnaN (нижняя панель). (E) Вестерн-блот-анализ клеточных лизатов указанных генотипов, выращенных с ксилозой или без нее, с использованием антисыворотки против CtpA (верхняя панель) и DnaN (нижняя панель).
https://doi.org/10.1371/journal.pgen.1007512.g002
YlbL и CtpA имеют перекрывающиеся функции
Сходство фенотипов привело нас к гипотезе, что YlbL и CtpA имеют перекрывающиеся функции. Чтобы проверить это, мы выполнили эксперимент по кросс-комплементации, используя точечный анализ титра чувствительности MMC.Сверхэкспрессия YlbL, но не YlbL-S234A, дополняла делецию ctpA (фиг. 3A и 3B). Точно так же сверхэкспрессия CtpA, но не CtpA-S297A, дополняла делецию ylbL (фиг. 3A и 3B). Кроме того, делеция обеих протеаз превратила B в . subtilis гиперчувствительность к MMC, даже в большей степени, чем потеря uvrA , который кодирует белок, ответственный за распознавание аддуктов MMC как часть эксцизионной репарации нуклеотидов [Рис. 3C; 38, 39].Чтобы дополнительно проверить гипотезу о том, что YlbL и CtpA имеют перекрывающиеся функции, мы сверхэкспрессировали каждую из протеаз отдельно на фоне двойных мутантных протеаз и наблюдали полное восстановление чувствительности MMC при экспрессии дикого типа (WT), но не серина. варианты (Рис. 3D и 3E).
Рис. 3. YlbL и CtpA имеют перекрывающиеся функции.
(A) Анализ точечного титра с использованием указанных генотипов и сред. (B) Вестерн-блот-анализ клеточных лизатов генотипов на панели A с использованием указанной антисыворотки. (C&D) Анализ точечного титра с использованием указанных генотипов и сред. (E) Вестерн-блот-анализ клеточных лизатов генотипов, указанных на панели D, с использованием указанной антисыворотки.
https://doi.org/10.1371/journal.pgen.1007512.g003
Задержка деления клеток, зависящая от повреждения ДНК, увеличивается при делециях протеазы
Эксперименты, проведенные до сих пор, не позволяют различить чувствительность к MMC в результате гибели клеток, ингибирования роста или того и другого.Чтобы определить, возникает ли чувствительность из-за гибели клеток, мы провели анализ выживаемости с использованием острого лечения MMC. Мы обнаружили небольшое снижение процента выживаемости по мере увеличения концентрации ММС в Δ ylbL и штамме с двойным мутантом (S4A фиг.). Мы сравнили снижение процента выживаемости у одинарных и двойных мутантов протеазы со штаммом Δ uvrA , который, как было показано ранее, очень чувствителен к MMC [40]. Штамм, лишенный uvrA , был очень чувствителен к острому лечению MMC (S4A фиг.), Тогда как штамм с двойной делецией протеазы был значительно менее чувствителен к острому воздействию по сравнению с Δ uvrA (сравните фиг. 3C и S4A).Взятые вместе, мы пришли к выводу, что чувствительность ММС мутантов протеазы, наблюдаемая в анализах титров пятен, в первую очередь вызвана ингибированием роста.
Мы предположили, что чувствительность к повреждению ДНК в результате ингибирования роста также может быть объяснена ингибированием пролиферации клеток или ингибированием деления клеток, а не роста клеток. Чтобы различать эти две возможности, мы измерили длину клетки, потому что ингибирование пролиферации должно наблюдаться как увеличение длины клетки, что согласуется с неудачей в восстановлении контрольной точки.Таким образом, мы разработали анализ восстановления MMC, аргументируя это тем, что после обработки MMC клетки, лишенные YlbL, CtpA или обоих, останутся удлиненными, показывая более медленное восстановление контрольной точки по сравнению со штаммом WT. Мы выращивали культуры либо в контрольном носителе, либо в присутствии MMC. После двухчасовой обработки среду, содержащую MMC, удаляли и клетки промывали. Затем клетки переносили в свежую среду без MMC и позволяли продолжить рост для анализа восстановления контрольной точки. Хотя клетки оказались удлиненными в штаммах Δ ylbL и двойных мутантных штаммах, в популяции наблюдалась гетерогенность (фиг. 4A).В результате мы измерили длину не менее 900 клеток для каждого генотипа и каждого состояния и построили распределение длины клеток в виде гистограмм (рис. 4B). Не было никакой разницы в распределении длины ячейки управления транспортным средством (рис. 4B). Обработка всех штаммов MMC привела к сдвигу вправо в распределении для всех штаммов (рис. 4B, сравните верхние панели). При сравнении делеций протеазы с WT разницу в распределении можно было визуализировать, рассматривая процент клеток больше 6.75 мкм, что составляет примерно три ячейки длиной 2,25 мкм каждая. Мы обнаружили, что делеция ylbL приводила к увеличению процента клеток длиной более 6,75 мкм в культурах, обработанных MMC, и после двух и четырех часов восстановления (рис. 4C). Однако удаление ctpA привело к очень небольшому, хотя и значительному (p-значение = 0,0142 для одностороннего Z-теста) увеличению процента клеток длиной более 6,75 мкм после 4 часов восстановления (рис. 4C). Двойной мутант привел к тому, что процент клеток был немного больше, чем только Δ ylbL через два часа (p-значение = 0.0001 для одностороннего Z-теста) и четыре часа (p-значение = 0,0088 для одностороннего Z-теста) восстановления (рис. 4C).
Рис. 4. Повреждение ДНК задерживает цитокинез в клетках с делециями протеазы.
(A) Типичные микрофотографии клеток с указанными генотипами в указанные моменты времени. Мембраны окрашивали FM4-64. Масштабная линейка 5 мкм. (B) Распределение длины клеток в виде гистограмм. Ось ординат на всех графиках нормирована на размер образца, дающий плотность, а ось абсцисс — длина ячейки в мкм.Количество клеток, оцененных в каждом распределении, указано как «n =», и указан генотип каждого штамма. Пунктирная вертикальная линия в распределении «носитель» нанесена на 2,25 мкм, это приблизительное среднее значение для всех штаммов. Пунктирная вертикальная линия в остальных распределениях составляет 6,75 мкм, что в три раза превышает среднюю длину необработанных клеток. (C) Доля клеток, представленных гистограммами на панели B с длиной более 6,75 мкм, представлена в виде гистограммы.Планки погрешностей представляют собой стандартное отклонение, где p представляет собой долю, а n — размер выборки. Звездочка указывает значение p менее 0,05.
https://doi.org/10.1371/journal.pgen.1007512.g004
В совокупности мы заключаем, что YlbL является первичной протеазой в этих условиях, и CtpA также вносит свой вклад. Мы также пришли к выводу, что клеткам, лишенным YlbL или одновременно YlbL и CtpA, требуется больше времени для деления после воздействия MMC, что согласуется с чувствительностью к повреждению ДНК в результате ингибирования пролиферации клеток.Кроме того, наблюдение за ингибированием пролиферации клеток предполагает, что протеазы YlbL и CtpA могут быть важны для восстановления контрольной точки повреждения ДНК (см. Ниже).
Уровни YlbL и CtpA не регулируются повреждением ДНК
Потенциальной моделью для регулирования YlbL и CtpA в ответ на повреждение ДНК является повышение уровня белка после воздействия повреждения ДНК. Повышенные уровни протеазы в ответ на повреждение ДНК могут способствовать восстановлению контрольной точки повреждения ДНК, когда это необходимо. Чтобы проверить эту модель, мы контролировали уровни белков YlbL и CtpA с помощью вестерн-блоттинга в ходе анализа восстановления MMC.Уровни белка YlbL и CtpA не изменялись относительно загружающего контрольного DnaN на протяжении всего эксперимента (S4B и S4C фиг.). В качестве положительного контроля мы провели тот же эксперимент и отслеживали уровни белка RecA и обнаружили, что действительно уровни белка RecA увеличились (S4B и S4C, рис.), Как и ожидалось, поскольку recA индуцируется как часть SOS-ответа [42, 43 ]. Мы пришли к выводу, что уровни белков YlbL и CtpA не регулируются повреждением ДНК.
Ингибитор клеточного деления YneA накапливается в мутантах протеазы
Данные, представленные до сих пор, привели нас к предположению, что в отсутствие YlbL и CtpA белок накапливается, что приводит к ингибированию клеточного деления (рис. 5A).Для выявления накапливающегося белка мы провели анализ всего протеома экстрактов клеток дикого типа и двойных мутантных протеаз. Мы решили проанализировать протеомы клеток после двух часов восстановления, потому что распределения длины клеток больше всего различались между WT и двойным мутантом протеазы (рис. 4B). Мы обнаружили, что нормализованные данные спектрального подсчета имели сходные распределения как для дикого животного, так и для двойного мутанта, которые были приблизительно логарифмически нормальными (S5A фиг.). Мы проверили, что распределение тестовой статистики (разница в среднем двойном мутанте и среднем WT) было нормально распределено (S5B Рис.), Что позволяет использовать t-критерий.Мы также выполнили анализ основных компонентов и обнаружили, что WT реплицируется и двойной мутант реплицируется, каждый сгруппирован вместе (S5C фиг.).
Рис. 5. YneA накапливается в мутантах протеазы.
(A) Модель функции YlbL и CtpA в регуляции деления клеток. (B) Данные протеомики представлены как кратное изменение (двойной мутант / WT) в зависимости от p-значения. Точки, отмеченные черным цветом, имеют кратное изменение меньше единицы или значение p, большее или равное 0,05, а точки, нанесенные на график красным цветом, имеют кратное изменение больше единицы и значение p меньше 0.05. (C) Вестерн-блоттинг анализ клеточных лизатов из штаммов с указанными генотипами в указанные моменты времени из анализа выделения MMC (см. Методы) с использованием антисыворотки к YneA или DnaN.
https://doi.org/10.1371/journal.pgen.1007512.g005
Всего было обнаружено 2329 белков (таблица S4), и было обнаружено, что 183 белка дифференциально представлены (значение p <0,05) в двойном мутант по отношению к WT (таблица S5). Из белков, дифференциально представленных в двойном мутанте, 104 имели кратное изменение больше единицы (фиг. 5B, красные точки).В B описаны три основных механизма. subtilis для ингибирования деления клеток: 1) Noc-зависимая окклюзия нуклеоидов [44], 2) истощение FtsL [45, 46] и 3) экспрессия YneA [23]. Одна возможность заключалась в том, что уровни белка Noc были выше у двойного мутанта, но мы не наблюдали разницы в уровнях Noc (S5D, фиг.). Другая возможность заключалась в том, что FtsL или протеаза RasP, которая деградирует FtsL, была затронута на фоне мутантной протеазы [46]. Мы не обнаружили различий в относительном содержании белка FtsL или RasP (S5D фиг.), Что исключает путь FtsL / RasP.Среди 10 лучших белков, которые были более многочисленны в двойном мутанте, был YneA, SOS-зависимый ингибитор деления клеток (таблица S5). Мы спросили, произошло ли обогащение YneA просто потому, что оно вызвано SOS. Мы проанализировали относительное содержание нескольких других белков, которые, как известно, индуцируются SOS, включая RecA, UvrA, UvrB, DinB и YneB [42], который находится в опероне с YneA [23]. Мы обнаружили, что ни один из этих других белков не был обогащен двойным мутантом (S5E фиг.). Эти результаты предполагают, что накопление YneA не является результатом повышенной активации SOS, и регуляция накопления YneA, вероятно, будет посттрансляционной, потому что уровни белка другого члена оперона, YneB, не изменились.Взятые вместе, наши протеомические данные предполагают, что YlbL и CtpA способствуют восстановлению контрольных точек повреждения ДНК посредством регулирования количества белка YneA.
Мы непосредственно тестировали накопление YneA в мутантах протеазы на протяжении всего анализа выделения MMC с помощью вестерн-блоттинга. YneA накапливался во всех штаммах с делецией протеазы через 2 и 4 часа восстановления, хотя накопление YneA в Δ ctpA было небольшим (фиг. 5C). В двойном мутанте YneA накапливался в условиях обработки MMC в дополнение к обеим точкам времени восстановления (рис. 5C).В двойном мутанте мы наблюдали несколько видов YneA, что, как мы предполагаем, является результатом неестественно высоких уровней белка YneA, приводящих к неспецифическому расщеплению другими протеазами. С этими результатами мы предполагаем, что YneA является субстратом YlbL и CtpA, оба из которых разрушают YneA, обеспечивая восстановление контрольной точки.
yneA требуется для фенотипов чувствительности к повреждению ДНК и удлинения клетокХотя накопление YneA хорошо согласуется с нашими данными, мы сочли, что другие белки, обогащенные более чем в пять раз двойным мутантом, могли вносить вклад в фенотип чувствительности к повреждению ДНК.Чтобы проверить это, мы сконструировали делеции каждого гена в WT и двойном мутанте и проверили чувствительность MMC. Мы обнаружили, что ни одна единственная делеция каждого из 10 генов не приводила к чувствительности к MMC (S6A фиг.). В двойном мутанте только делеция yneA была способна восстановить чувствительность к MMC (S6A фиг.). Мы подтвердили, что делеция yneA может спасти чувствительность MMC во всех фонах мутантных протеаз (рис. 6A). Мы исследовали длину клетки в анализе восстановления повреждений ДНК.Как и ожидалось, делеция yneA приводила к менее серьезному удлинению клеток по сравнению с WT (сравните WT на фиг. 4B и Δ yneA∷loxP на фиг. 6B). Кроме того, делеция ylbL , ctpA или обоих больше не изменяла распределение длины клеток в отсутствие yneA в двухчасовой точке времени восстановления (Фиг.6B, 6C и S6B). При лечении MMC мы действительно наблюдали небольшое увеличение (значение p = 0,0004 для одностороннего Z-теста) процента клеток больше 6.75 мкм в штамме с двойной делецией протеазы по сравнению с WT (фиг. 6C). Учитывая, что чувствительность MMC и большая часть удлинения клеток у мутантов протеазы зависит от yneA , мы предположили, что экспрессии только YneA будет достаточно для подавления роста мутантов протеазы в большей степени. Действительно, штаммы, лишенные YlbL, CtpA или обоих, были более чувствительны к сверхэкспрессии yneA с индуцибельного промотора IPTG, чем WT (фиг. 6D). Кроме того, мы показываем, что YneA накапливается в штаммах мутантных протеаз после эктопической экспрессии yneA (рис. 6E).Мы пришли к выводу, что накопление YneA приводит к серьезному ингибированию роста клеток, лишенных YlbL и CtpA.
Рис. 6. yneA необходим для определения чувствительности к повреждению ДНК и фенотипов удлинения клеток.
(A) Анализ точечного титра с использованием указанных генотипов и сред. (B) Распределение длины клеток в виде гистограмм. Количество клеток, оцененных в каждом распределении, указано как «n =», а генотип каждого штамма указан над распределениями.Пунктирная вертикальная линия в распределении «Транспортное средство» соответствует приблизительному среднему значению 2,25 мкм. Пунктирная вертикальная линия в остальных распределениях соответствует 6,75 мкм. Ось ординат на всех графиках нормирована на размер образца, дающий плотность, а ось абсцисс — длина ячейки в мкм. (C) Доля клеток размером более 6,75 мкм из распределений на панели B представлена в виде гистограммы. Планки погрешностей представляют собой стандартное отклонение, где p представляет собой долю, а n — размер выборки.Звездочка указывает значение p менее 0,05. (D) Точечный анализ титра, проверяющий эффект избыточной продукции YneA с использованием указанной концентрации индуктора IPTG. (E) Вестерн-блот-анализ клеточных лизатов с использованием 100 мкМ IPTG для экспрессии YneA с указанными генотипами с использованием указанной антисыворотки.
https://doi.org/10.1371/journal.pgen.1007512.g006
CtpA специфически переваривает YneA
in vitroЧтобы проверить гипотезу о том, что YneA является прямым субстратом протеаз, мы очистили YneA (a.а. 28–103), CtpA (а.о. 38–466) и YlbL (а.о. 36–341), лишенные своих N-концевых трансмембранных доменов, чтобы сделать возможным изоляцию. Нам не удалось обнаружить протеазную активность из YlbL с использованием YneA, лизоцима или казеина в качестве субстратов (S7 Рис; см. Обсуждение). Когда очищенный CtpA инкубировали с YneA, мы наблюдали переваривание YneA с течением времени, но не наблюдали переваривания с использованием CtpA-S297A (фиг. 7A). Чтобы проверить, является ли активность CtpA против YneA специфической, мы завершили ту же реакцию, используя лизоцим в качестве субстрата, и не обнаружили активности (рис. 7B).Мы пришли к выводу, что YneA является прямым и специфическим субстратом CtpA.
Рис. 7. Восстановление контрольной точки повреждения ДНК в Bacillus subtilis .
(A) Анализ протеазы, инкубирующий очищенный CtpA или CtpA-S297A с очищенным YneA в течение указанного времени с последующим SDS-PAGE и окрашиванием кумасси синим. (B) Протеазный анализ, инкубирующий очищенный CtpA или CtpA-S297A с коммерчески доступным лизоцимом в течение 3 часов с последующим SDS-PAGE и окрашиванием кумасси синим. (C) Бактериальный двухгибридный анализ с использованием слияния T18 (строки) и слияния T25 (столбцы), котрансформированного в E . coli . Плазмидами T25 являются: пустой вектор (T25), T25-YlbL-S234A (T25-YlbL) и T25-CtpA-S297A (T25-CtpA). Плазмидами T18 являются: пустой вектор (T18), T18-YneA и T18-YneAΔN (в котором отсутствует трансмембранный домен). (D) Модель восстановления YlbL- и CtpA-зависимых контрольных точек повреждений ДНК. YlbL и CtpA присутствуют в виде мембранных протеаз, и когда присутствует большое количество повреждений ДНК, продукция YneA подавляет обе протеазы, что приводит к задержке деления клеток.После завершения репарации ДНК и снижения экспрессии YneA, YneA очищается и клеточное деление продолжается.
https://doi.org/10.1371/journal.pgen.1007512.g007
YlbL-S234A и CtpA-S297A взаимодействуют с YneA
Хотя мы не смогли обнаружить активность протеазы YlbL in vitro , мы спросили, может ли YlbL взаимодействовать с YneA. Чтобы проверить это, мы использовали бактериальный двугибридный анализ [47, 48]. Мы использовали этот анализ, потому что он эффективен при обнаружении взаимодействий между мембранными белками [22, 26, 49].Мы протестировали YlbL-S234A и CtpA-S297A для предотвращения переваривания YneA и проанализировали взаимодействие с полноразмерным YneA или YneA без его трансмембранного домена (YneAΔN) в качестве контроля. Мы обнаружили, что YlbL и CtpA взаимодействовали с полноразмерным YneA (рис. 7C), но не было обнаружено взаимодействия с YneAΔN (рис. 7C), вероятно, из-за того, что YneA не локализовался на мембране. Учитывая, что YlbL не обладал активностью in vitro и мы обнаружили взаимодействие с YneA в бактериальном двухгибридном анализе, мы предполагаем, что YneA является прямым субстратом YlbL и что YlbL требует полноразмерного YneA для взаимодействия.
Обсуждение
Как YlbL и CtpA распознают YneA в качестве субстрата? Интригующий аспект этого механизма восстановления контрольных точек — использование неродственных протеаз. И YlbL, и CtpA имеют трансмембранные домены и домены PDZ, но домены пептидазы сильно различаются. CtpA имеет домен S41 пептидазы и гомологичен Tail-специфической протеазе или Tsp (также Prc), которая распознает С-конец своего субстрата через его домен PDZ [50, 51]. Мы предполагаем, что CtpA также распознает YneA через домен PDZ и что этот механизм объясняет, как CtpA распознает другие родственные ему субстраты.Фактически, исследование Мо и Буркхолдера идентифицировало остаток на С-конце YneA (D97), который при мутации в аланин стабилизирует YneA [27]. Возникает соблазн предположить, что D97 в YneA важен для CtpA для распознавания YneA. Механизм, с помощью которого YlbL распознает YneA, менее ясен. YlbL имеет уникальную доменную организацию, не обнаруженную в других изученных протеазах. YlbL не имеет домена AAA + ATPase, общего для других протеаз Lon, что логично, учитывая, что YlbL, вероятно, находится вне клетки и разрушает YneA.Вместо домена АТФазы YlbL имеет домен PDZ, который может действовать как домен распознавания субстрата или как ингибирующий домен, подобный домену PDZ DegS [52-54]. Бактериальный двухгибридный анализ предполагает, что YlbL действительно распознает YneA напрямую (рис. 7C), хотя мы не можем исключить возможность того, что существует адаптерный белок, который распознает YneA, когда эти белки привязаны к мембране. Мы также не идентифицировали потенциальный адаптер в данных Tn-seq, что позволяет предположить, что прямое взаимодействие действительно происходит между YlbL и YneA.Последняя возможность состоит в том, что YlbL распознает YneA через трансмембранный домен, который затем активирует домен пептидазы Lon для разрушения или расщепления YneA. Это также могло бы объяснить причину, по которой мы не смогли обнаружить активность с использованием YlbL, у которого отсутствует его трансмембранный домен. В любом случае необходимы дальнейшие эксперименты, чтобы выяснить механизм, с помощью которого YlbL узнает YneA.
Все организмы контролируют клеточные процессы посредством регулируемой передачи сигналов. Чтобы регулировать клеточный процесс, сигнальный путь должен иметь механизмы активации и инактивации.Многие бактерии используют небольшой мембранный белок в качестве белка контрольной точки повреждения ДНК, вызванного SOS [22-25]. Однако механизм восстановления контрольных точек для организмов, использующих контрольные точки мембранного белка, остается неясным. Наше всестороннее исследование выявило двойной протеазный механизм восстановления контрольной точки повреждения ДНК (рис. 7D). Протеазы YlbL и CtpA постоянно присутствуют в плазматической мембране клеток даже при отсутствии повреждений ДНК. После обнаружения повреждения ДНК индуцируется экспрессия YneA. Мы предполагаем, что активности YlbL и CtpA насыщаются за счет увеличения экспрессии YneA, что приводит к задержке клеточного деления.После репарации ДНК экспрессия YneA снижается, а YlbL и CtpA очищают любой оставшийся YneA, позволяя возобновить деление клеток.
Контрольные точки повреждения ДНКимеют фундаментальное значение для биологии, и мы обнаружили путь, ответственный за инактивацию контрольных точек и повторное включение клеточного цикла в B . subtilis . Эти результаты представляют собой важный шаг вперед в определении того, как происходит восстановление контрольных точек у бактерий. Ингибиторы мембраносвязанного деления клеток, идентифицированные к настоящему времени [22–25], не являются гомологами YneA, и фактически единственной объединяющей особенностью является то, что они являются небольшими мембраносвязанными белками [22–25].Это представляет собой серьезную проблему, потому что компоненты обеспечения соблюдения контрольных точек и восстановления должны быть определены экспериментально. Наше исследование служит моделью для идентификации протеаз восстановления контрольных точек с помощью прямой генетики, которая, в свою очередь, может быть использована для идентификации белка принудительного применения с помощью протеомики. Важнейшей особенностью нашего подхода было использование нескольких периодов роста в Tn-seq, что позволило нам идентифицировать обе протеазы. Таким образом, мы предлагаем стратегию, использующую комбинированные подходы прямой генетики и целевой протеомики для идентификации путей контрольных точек повреждения ДНК в генетически трактуемых бактериальных патогенах.
Восстановление после контрольной точки повреждения ДНК — критический процесс для всех организмов. Одна тема, встречающаяся во всей биологии, — это использование нескольких белков с перекрывающимися функциями. У эукариот события фосфорилирования, которые устанавливают контрольную точку, удаляются множественными фосфатазами [55, 56]. В E . coli , есть две цитоплазматические протеазы, Lon и ClpYQ, которые, как было обнаружено, разрушают ингибитор клеточного деления SulA [16, 18–20]. Наше исследование дополнительно расширяет использование нескольких факторов в регулировании восстановления контрольных точек до B . subtilis , описывая механизм с использованием двух протеаз. У эукариот множественные белки с перекрывающимися функциями часто существуют из-за пространственных или временных ограничений, которые, по-видимому, частично объясняют использование множественных факторов в восстановлении контрольных точек [55, 56]. В E . coli , ClpYQ оказался важным при более высоких температурах в отсутствие Lon [19], что снова указывает на то, что каждая протеаза функционирует в определенных условиях. В случае с YlbL и CtpA, однако, в мультимедийных материалах, по всей видимости, лежит общая ответственность.Удаление каждой протеазы приводит к чувствительности к повреждению ДНК, а двойной мутант имеет более серьезную чувствительность. Напротив, во время роста в минимальной среде YlbL, по-видимому, является первичной протеазой, поскольку фенотип клеточного удлинения более выражен в клетках, лишенных ylbL . До сих пор неясно, как и когда функционирует каждая протеаза. Почему для разложения YneA недостаточно одной протеазы? Занимают ли протеазы отдельные локусы в клетке, требуя, чтобы каждая протеаза деградировала определенный пул YneA? Другая возможность заключается в том, что уровни протеазы ограничены другим эволюционным давлением, например субстратами, уникальными для каждой протеазы.Таким образом, клетка не может поддерживать отдельные протеазы на уровнях, необходимых для титрования YneA как части DDR, потому что уровни другого субстрата будут слишком низкими. Другое объяснение состоит в том, что использование нескольких факторов — это эволюционная стратегия, которая увеличивает приспособленность организма. Ясно, что восстановление контрольной точки имеет решающее значение, потому что приспособленность клеток, лишенных ylbL или ctpA , значительно снижается при наличии повреждения ДНК (таблица S2).
Хотя описанный здесь механизм двойной протеазы решает важный шаг в DDR, наши данные также показывают сложность системы.После того, как мы подвергали клетки воздействию MMC, клетки удлинялись. Однако мы заметили, что не вся элонгация зависит от yneA (см. Фиг. 6B), что позволяет предположить другой механизм контроля клеточного цикла. В B . subtilis , были сообщения о yneA -независимом контроле клеточного деления после стресса репликации [45, 57, 58]. Важный компонент клеточного деления FtsL, как сообщается, нестабилен, и его истощение ведет к ингибированию клеточного деления [58]. Кроме того, сообщалось, что уровни транскрипта ftsL уменьшаются после стресса репликации независимо от SOS-ответа [45], таким образом связывая истощение нестабильного белка FtsL с контролем клеточного деления после стресса репликации.В исследовании с использованием блока репликации, состоящего из Tet-репрессора, связанного с массивом Tet-оператора, наблюдалось ингибирование деления клеток, не зависящее от yneA , noc и FtsL [57]. Интересно, что недавние исследования Caulobacter crescentus выявили два ингибитора клеточного деления, которые экспрессируются в ответ на повреждение ДНК, причем один ингибитор SOS-зависимый, а другой SOS-независимый [22, 26]. В B . megaterium , недавнее исследование показало, что транскрипт yneA нестабилен после воздействия повреждения ДНК [59], что указывает на еще один уровень регуляции.Фактор, регулирующий транскрипты yneA в предыдущем исследовании, не был идентифицирован, хотя возможно, что один из генов с неизвестной функцией, идентифицированный в наших скринингах, мог регулировать мРНК yneA . Вместе эти исследования подчеркивают сложность регулирования контрольной точки повреждения ДНК у бактерий.
Материалы и методы
Бактериологические методы и химические препараты
Бактериальные штаммы, плазмиды и олигонуклеотиды, использованные в этом исследовании, перечислены в таблице S6, а создание штаммов и плазмид подробно описано в дополнительных методах.Все штаммы Bacillus subtilis являются изогенными производными PY79 [60]. Bacillus subtilis Штаммы выращивали в LB (10 г / л NaCl, 10 г / л триптона и 5 г / л дрожжевого экстракта) или минимальной среде S7 50 с 2% глюкозы (1x S7 50 солей (разбавленных из 10x S7 50 солей: 104,7 г / л MOPS, 13,2 г / л, сульфат аммония, 6,8 г / л одноосновного фосфата калия, pH 7,0, скорректированный гидроксидом калия), 1x металлов (разбавленный из 100x металлов: 0,2 M MgCl 2 , 70 мМ CaCl 2 , 5 мМ MnCl 2 , 0.1 мМ ZnCl 2 , 100 мкг / мл тиамин-HCl, 2 мМ HCl, 0,5 мМ FeCl ( 3 ), 0,1% глутамата калия, 2% глюкозы, 40 мкг / мл фенилаланина, 40 мкг / мл триптофана) при 30 ° C при встряхивании (200 об / мин). Митомицин С (MMC), метилметансульфонат (MMS) и флеомицин использовали в концентрациях, указанных на фигурах. Следующие антибиотики были использованы для селекции в B . subtilis , как указано в деталях метода: спектиномицин (100 мкг / мл), хлорамфеникол (5 мкг / мл) и эритромицин (0.5 мкг / мл). Отбор трансформантов Escherichia coli (клетки MC1061 или TOP10 для клонирования или BL21 для экспрессии белка) проводили с использованием следующих антибиотиков: спектиномицин (100 мкг / мл) или канамицин (50 мкг / мл).
Tn-seq
Библиотека вставки транспозонов была сконструирована аналогично [61] с модификациями, описанными в дополнительных методах. Эксперименты Tn-seq были разработаны с несколькими периодами роста, подобными предыдущему описанию [28], с подробным описанием в дополнительных методах.Создание библиотеки секвенирования и анализ данных выполняли, как описано ранее [35, 61], с модификациями, описанными в дополнительных методах. Данные секвенирования были депонированы в базу данных GEO с регистрационным номером GSE109366.
Точечный анализ титра
Б . Штаммы subtilis высевали на агар LB и инкубировали при 30 ° C в течение ночи. На следующий день одну колонию использовали для инокуляции 2 мл культуры LB в 14 мл круглодонной культуральной пробирке, которую инкубировали при 37 ° C на вращающейся штативе до тех пор, пока OD 600 не станет равным 0.5–1. Культуры нормализовали до OD 900 = 0,5 и серийно разводили. Серийные разведения наносили каплями (4 мкл) на агаровую среду, указанную на рисунках, и планшеты инкубировали при 30 ° C в течение ночи (16–20 часов). Все точечные анализы титра проводили как минимум дважды.
Анализы на выживаемость
Анализы выживаемости с использованием экстренной терапии митомицином С выполняли, как описано ранее [62]. Культуры выращивали до OD 900 , равного примерно 1, и трех образцов, равных 0.Брали 6 мл OD 600 = 1 эквивалент, и клетки осаждали центрифугированием: 10 000 г в течение 5 минут при комнатной температуре (все последующие стадии центрифугирования были идентичными). Клетки промывали 0,6 мл 0,85% NaCl (физиологический раствор) и осаждали центрифугированием. Осадки клеток ресуспендировали в 0,6 мл физиологического раствора и аликвоты по 100 мкл распределяли для каждой концентрации ММС. MMC добавляли в каждую пробирку для получения конечной концентрации, указанной на рисунке, в общем объеме 200 мкл, и клетки инкубировали при 37 ° C в течение 30 минут.Клетки осаждали центрифугированием для удаления ММС, ресуспендировали в физиологическом растворе и серийное разведение, дающее поддающееся оценке количество клеток (около 30–300), высевали на агар LB для определения выживающей фракции клеток. Каждый эксперимент проводился трижды в трех экземплярах для каждого штамма.
Производство антисыворотки
Очищенные белки (см. Дополнительные методы для протоколов очистки) были отправлены в Covance для получения антител с использованием кроликов. В 77-дневном протоколе использовали двух кроликов, а для вестерн-блоттинга использовали сыворотку с наименьшим фоном.
Вестерн-блоттинг
Для вестерн-блоттинга YlbL, CtpA, RecA и DnaN осадок клеток, эквивалентный 1 мл OD 600 = 1, ресуспендировали в 100 мкл 1x буфера SMM (0,5 M сахароза, 0,02 M малеиновая кислота, 0,02 M MgCl 2 , доведенный до pH 6,5), содержащий 1 мг / мл лизоцима и 2x ингибитора протеазы Roche, при комнатной температуре в течение 1 или 2 часов. Затем образцы лизировали добавлением 6x загрузочного красителя SDS (0,35 M Трис, pH 6,8, 30% глицерина, 10% SDS, 0,6 M DTT и 0,012% бромфенолового синего) к 1x.Образцы (12 мкл) разделяли с помощью 10% SDS-PAGE и переносили на нитроцеллюлозу с помощью Trans-Blot Turbo (BioRad) в соответствии с инструкциями производителя. Мембраны блокировали в 5% молоке в TBST (25 мМ Трис, pH 7,5, 150 мМ NaCl и 0,1% Твин 20) при комнатной температуре в течение 1 часа или при 4 ° C в течение ночи. Блокирующий буфер удаляли, и первичные антитела добавляли в 2% молоко в TBST (αYlbL, 1: 5000 или 1: 8000; αCtpA, 1: 5000; αRecA, 1: 4000; αDnaN, 1: 4000). Инкубацию первичных антител проводили при комнатной температуре в течение 1 часа или в течение ночи при 4 ° C.Первичные антитела удаляли, и мембраны трижды промывали TBST в течение 5 минут при комнатной температуре. Вторичные антитела (Licor; 1: 15000) добавляли в 2% молоко в TBST и инкубировали при комнатной температуре в течение 1 часа. Мембраны трижды промывали, как указано выше, и визуализировали с помощью системы визуализации Li-COR Odyssey. Все эксперименты вестерн-блоттинга проводились как минимум дважды с независимыми образцами. Маркеры молекулярной массы использовали в блотах YlbL и CtpA. YlbL мигрирует в место между маркерами 30 и 40 кДа, что соответствует его предсказанной молекулярной массе 37.6 кДа. CtpA мигрировал в место между маркерами 40 и 80 кДа, что соответствовало его молекулярной массе 51,1 кДа.
Для вестерн-блоттинга YneA осадки клеток, 10 мл OD 600 = 1 для анализа выделения MMC и 25 мл OD 900 = 1 для сверхэкспрессии, ресуспендировали в 400 или 500 мкл, соответственно, буфера для обработки ультразвуком. (50 мМ Трис, pH 8,0, 10 мМ EDTA, 20% глицерин, 2x ингибиторы протеазы Roche и 5 мМ PMSF) и лизировали с помощью обработки ультразвуком. Загрузочный краситель SDS добавляли в 2 раза и образцы инкубировали при 100 ° C в течение 7 минут.Образцы (10 мкл) разделяли с использованием 16,5% Tris-Tricine-SDS-PAGE (BioRad) и переносили на нитроцеллюлозную мембрану с использованием Trans-blot Turbo (BioRad) в соответствии с инструкциями производителя. Все последующие этапы были выполнены, как указано выше, с разведением первичных антител 1: 3000.
Анализ выделения митомицина С
Планшет с агаром LB, выращенный при 30 ° C в течение ночи, промывали предварительно нагретой минимальной средой S7 50 и использовали для инокуляции культуры минимальной среды S7 50 при OD 600 = 0.1. Культуры инкубировали при 30 ° C до OD 600 около 0,2 (2–2,5 часа). MMC добавляли до 100 нг / мл и культуры инкубировали при 30 ° C в течение 2 часов. Клетки осаждали центрифугированием (4696 г в течение 7 минут) и среду удаляли. Осадки клеток промывали равным объемом 1x PBS, pH 7,4 и снова осаждали центрифугированием, как описано выше. Осадки клеток ресуспендировали в равном объеме предварительно нагретой минимальной среды S7 50 и инкубировали при 30 ° C в течение четырех часов.Образцы для микроскопии и вестерн-блоттинга отбирали после двухчасовой обработки ММС и через два и четыре часа после восстановления, как показано на фигурах. Контрольные образцы носителя обрабатывали в течение 2 часов эквивалентным объемом носителя, в котором была суспендирована ММС (25% (об. / Об.) ДМСО).
Микроскопия
Был взят образец объемом 500 мкл из анализа выделения MMC, описанный выше, и FM4-64 был добавлен до 2 мкг / мл и инкубирован при комнатной температуре в течение 5 минут. Затем образцы переносили на 1% агарозные подушечки, сделанные из 1х солей Спизизена.Изображения были получены с помощью микроскопа Olympus BX61.
Анализ длины клеток
Клетки оценивали по длине клеток с помощью измерительного инструмента в программе ImageJ. Для каждого оцененного изображения были измерены все клетки, которые были в фокусе. Количество клеток, оцененных для каждого штамма / состояния, указано на рисунках (n = измеренные клетки). Гистограммы были созданы с использованием ggplot2 в R. Все оценки производились с использованием нескорректированных изображений. Репрезентативные изображения, показанные на рисунках, были изменены в ImageJ путем вычитания фона (метод радиуса катящегося шарика) и регулировки яркости и контрастности.Все внесенные изменения были применены ко всему изображению.
Детали эксперимента по протеомике
Образцы(5 мл OD 600 = 1) собирали из культур, выращенных, как описано в разделе анализа выделения MMC, при 2-часовом восстановлении центрифугированием: 4696 г при комнатной температуре в течение 10 минут. Образцы дважды промывали 500 мкл 1х PBS, pH 7,4 и осаждали центрифугированием: 10 000 г при комнатной температуре в течение 5 минут. Образцы замораживали в жидком азоте и хранили при -80 ° C.Образцы были отправлены на масс-спектрометрический анализ в MS Bioworks. Дальнейшая обработка образцов и анализ данных были выполнены MS Bioworks, как описано в дополнительных методах. Файлы необработанных данных доступны по запросу, а таблицы обработанных данных предоставляются в виде дополнительных таблиц (таблицы S4 и S5).
Анализы переваривания YneA и лизоцима
реакций расщепления YneA готовили в виде 20 мкл реакции в 20 мМ Трис pH 7,5, 20 мМ NaCl и 20% глицерине, содержащем 150 мкМ YneA и 2 мкМ CtpA.Реакции инкубировали при 30 ° C в течение времени, указанного на рисунке. Реакции останавливали добавлением 6x SDS-красителя к 1x и инкубированием при 100 ° C в течение 5 минут. Продукты реакции разделяли с помощью 16,5% трис-трицин SDS-PAGE. Белки определяли окрашиванием кумасси синим. Анализы переваривания лизоцима выполняли, как для YneA, с использованием лизоцима 2 мг / мл, и реакции инкубировали при 30 ° C в течение 3 часов.
Бактериальные двугибридные анализы
Бактериальный двухгибридный анализ выполняли, как описано ранее [47, 48, 63].Вкратце, гибридные плазмиды Т18 и Т25 котрансформировали в клетки BTh201, и котрансформанты были отобраны на агаре LB + 100 мкг / мл ампициллина + 50 мкг / мл канамицина при 37 ° C в течение ночи. Культуры LB + 100 мкг / мл ампициллина + 50 мкг / мл канамицина инокулировали с использованием нескольких колоний и инкубировали при 37 ° C в течение 90 минут. Культуры разбавляли в 250 раз и 4 мкл наносили на агар LB + 100 мкг / мл ампициллина + 50 мкг / мл канамицина + 0,5 мМ IPTG + 40 мкг / мл X-gal и инкубировали при 30 ° C в течение 48 часов, затем при комнатная температура в течение 24 часов.Яркость и контраст изображений были отрегулированы с помощью Adobe Photoshop, и изменения были применены ко всему изображению. Все бактериальные двугибридные анализы проводили как минимум дважды.
Вспомогательная информация
S1 Рис. Анализ данных Tn-seq.
(A) Распределения считывания секвенирования для мест вставки транспозона (Tn), содержащих более 0 считываний для каждой повторности образцов библиотеки (слева) или образцов заквасочной культуры (справа) из эксперимента MMC.Ось ординат — это частота Tn сайтов, а ось абсцисс — логарифм 10 считываний секвенирования. Пунктирная вертикальная линия проведена в log 10 (100). (B) Распределения считывания секвенирования нанесены на график для указанных образцов, как на панели A, за исключением того, что реплики были суммированы перед построением графика. Библиотека и закваски одинаковы на графиках Ctrl и MMC; Библиотека и заквасочные культуры одинаковы на графиках MMS и Phleo, которые показаны дважды в каждом случае для прямого сравнения.Пунктирная вертикальная линия проведена в log 10 (350). (C) Распределения относительной пригодности нанесены на график для вставок Tn с более чем 10 считываниями секвенирования в контрольных образцах. По оси ординат отложена частота появления Tn сайтов, а по оси абсцисс — относительная приспособленность. Пунктирная вертикальная линия проведена на уровне 1.0. Для указанных экспериментов нанесены все три периода роста.
https://doi.org/10.1371/journal.pgen.1007512.s001
(TIF)
S2 Рис. Идентификация каталитических остатков YlbL и CtpA.
(A) Lon-протеазный домен YlbL был выровнен с LonA и LonB из B . subtilis и Lon из E . coli . Выравнивания показывают, что YlbL содержит консервативную каталитическую диаду протеаз Lon, состоящую из серина 234 и лизина 279. (B) Домен протеазы S41 CtpA был выровнен с CtpB из B . subtilis и Prc из E . coli . Выравнивания показали, что CtpA содержит консервативную каталитическую триаду, состоящую из серина 297, лизина 322 и глутамина 326.
https://doi.org/10.1371/journal.pgen.1007512.s002
(TIF)
S3 Рис. Разрушение
ylbK приводит к полярному эффекту на ylbL .(A) Анализ точечного титра с использованием указанных генотипов и сред. (B) Схема локусов ylbK и ylbL с предполагаемым сайтом связывания рибосомы, предложенная для контроля трансляции ylbL , помечена желтым. (C) Вестерн-блот-анализ клеточных лизатов указанных генотипов с использованием антисыворотки к YlbL или DnaN.
https://doi.org/10.1371/journal.pgen.1007512.s003
(TIF)
S4 Рис. Уровни YlbL и CtpA не регулируются повреждением ДНК.
(A) Анализ выживаемости MMC с использованием штаммов с указанными генотипами для проверки, вызвана ли чувствительность MMC гибелью клеток. Концентрация ММС, используемая во время 30-минутной инкубации, указана на оси абсцисс, а ось ординат представляет собой процент клеток, выживших после обработки, относительно состояния отсутствия обработки (0 нг / мл).Каждая точка представляет собой среднее значение трех технических повторов из трех отдельных экспериментов (n = 9), а полосы ошибок представляют собой стандартную ошибку среднего. (B) Типичный вестерн-блот-анализ клеточных лизатов на протяжении всего анализа выделения MMC с использованием антисыворотки YlbL, CtpA, DnaN или RecA. (C) Количественная оценка данных вестерн-блоттинга в виде гистограммы. Столбики представляют собой среднее значение из трех экспериментов (YlbL, CtpA и DnaN) или двух экспериментов (RecA), а полосы ошибок представляют собой стандартное отклонение (YlbL и CtpA) или диапазон (RecA) измерений.По оси ординат отложены относительные уровни белка, которые представляют собой указанный уровень белка, нормализованный к контролю нагрузки, DnaN и измерению без обработки.
https://doi.org/10.1371/journal.pgen.1007512.s004
(TIF)
S5 Рис. YneA накапливается в мутантах протеазы.
(A) Средние значения нормализованных спектральных значений представлены в виде гистограмм для WT (синий) и Δ ylbL , Δ ctpA , двойной мутант (DM; красный). По оси ординат отложен счетчик, а по оси абсцисс — логарифм 10 (нормализованные спектральные счета для среднего значения трех повторов). (B) Распределение тестовой статистики (среднее значение WT – среднее значение DM) отображается в виде гистограммы. (C) Анализ основных компонентов был выполнен с использованием нормализованных спектральных отсчетов из образцов WT (синий) и DM (красный) с использованием функции «prcomp» в R. Первые две координаты нанесены на оси x и y. , соответственно. (D&E) Средние относительные уровни белка (WT / DM) из набора протеомных данных нанесены на график для указанных белков, а полосы ошибок представляют собой стандартное отклонение.На вставке к панели E для большей ясности показано более детальное изображение.
https://doi.org/10.1371/journal.pgen.1007512.s005
(TIF)
S6 Рис.
yneA требуется для определения чувствительности к повреждению ДНК и фенотипов удлинения клеток.(A) Штаммы с указанными генотипами (чашки слева) наносили на указанную среду (метки столбцов) и инкубировали при 30 ° C в течение ночи. Делеция yneA подавляет фенотип чувствительности к ММС с двойным мутантом (DM) Δ ylbL , Δ ctpA . (B) Типичные микрофотографии окрашенных FM4-64 клеток указанных генотипов в указанные моменты времени в анализе выделения MMC. Масштабная линейка составляет 5 мкм.
https://doi.org/10.1371/journal.pgen.1007512.s006
(TIF)
S7 Фиг. Очищенный YlbL без его N-концевой трансмембраны не обнаруживает протеазной активности
in vitro .(A) Анализ протеазы с использованием флуоресцентно меченного казеина. YlbL, YlbL-S234A, CtpA и CtpA-S297A, лишенные своих N-концевых трансмембранных доменов, инкубировали с казеином.Казеин был флуоресцентно помечен, так что сигнал гасился до тех пор, пока он не переваривается протеазой. (B) Анализ переваривания YneA. YlbL и CtpA инкубировали с возрастающими концентрациями YneA. Первая полоса — это маркер молекулярной массы (M). (C) Анализ переваривания лизоцима. YlbL инкубировали с лизоцимом.
https://doi.org/10.1371/journal.pgen.1007512.s007
(TIF)
S1 Таблица. Собранные данные Tn-seq.
OD 900 измерения, время инкубации, подсчет жизнеспособных клеток, скорость роста, оцененная на основе подсчета жизнеспособных клеток, количество поколений, оцененных на основе подсчета жизнеспособных клеток и времени инкубации, идентификаторов образцов секвенирования, считывания секвенирования, считывания нанесенного на карту и количества Tn вставок с более чем 10 чтениями для каждого образца.
https://doi.org/10.1371/journal.pgen.1007512.s008
(XLSX)
S2 Стол. Списки относительной пригодности tn-seq.
Относительные значения приспособленности для каждого гена с достаточными данными во всех трех периодах роста для всех трех экспериментов Tn-seq представлены вместе с скорректированным значением p (метод BH; см. Методы STAR). Перечисляются имена генов или теги локусов, а межгенные области помечаются как «ig» с номером.
https://doi.org/10.1371/journal.pgen.1007512.s009
(XLSX)
S3 Таблица. Список tn-seq перекрывается.
(Первая вкладка) Гены с наименьшей относительной приспособленностью с скорректированным значением p менее 0,01 перечислены для каждого эксперимента. (Вторая вкладка) Представлены перекрывающиеся гены во всех трех экспериментах с Tn-seq, а также перекрывающиеся гены при попарных сравнениях трех экспериментов с Tn-seq.
https://doi.org/10.1371/journal.pgen.1007512.s010
(XLSX)
Благодарности
Мы хотели бы поблагодарить сотрудников лаборатории Simmons за вдумчивые обсуждения на протяжении всей этой работы.Мы хотели бы поблагодарить доктора Джаякришнана Нандакумара за щедрый дар плазмид для сверхэкспрессии белка и доктора Дэвида Руднера за плазмиду pDR110.
Ссылки
- 1. Джексон С.П., Бартек Дж. Реакция на повреждение ДНК в биологии человека и болезнях. Природа. 2009. 461 (7267): 1071–8. Epub 2009/10/23. pmid: 19847258; Идентификатор PubMed Central PMCID: PMCPMC2
- 0.
- 2. Blanpain C, Mohrin M, Sotiropoulou PA, Passegue E. Ответ на повреждение ДНК в тканеспецифических и раковых стволовых клетках.Клеточная стволовая клетка. 2011; 8 (1): 16–29. Epub 2011/01/08. pmid: 21211780.
- 3. Kreuzer KN. Ответы на повреждение ДНК у прокариот: регулирование экспрессии генов, изменение моделей роста и манипулирование вилками репликации. Cold Spring Harb Perspect Biol. 2013; 5 (11): a012674. Epub 2013/10/08. pmid: 24097899; Идентификатор PubMed Central PMCID: PMCPMC3809575.
- 4. Мишель Б. После 30 лет исследований реакция бактерий на SOS все еще удивляет нас. PLoS биология. 2005; 3 (7): e255. Epub 2005/07/08.pmid: 16000023; Идентификатор PubMed Central PMCID: PMCPMC1174825.
- 5. Бахароглу З., Мазель Д. SOS, грозная стратегия бактерий против агрессии. Обзоры микробиологии FEMS. 2014. 38 (6): 1126–45. Epub 2014/06/14. pmid: 244.
- 6. Friedberg EC, Walker GC, Siede W., Wood RD, Schultz RA, Ellenberger T. Ремонт ДНК и мутагенез. 2-е изд. Вашингтон, округ Колумбия: ASM Press; 2006.
- 7. Санкар А., Линдси-Больц Л.А., Унсал-Качмаз К., Линн С. Молекулярные механизмы репарации ДНК млекопитающих и контрольные точки повреждения ДНК.Ежегодный обзор биохимии. 2004. 73: 39–85. Epub 2004/06/11. pmid: 15189136.
- 8. Ciccia A, Elledge SJ. Реакция на повреждение ДНК: безопасная игра с ножами. Mol Cell. 2010. 40 (2): 179–204. Epub 2010/10/23. pmid: 20965415; Идентификатор PubMed Central PMCID: PMCPMC2988877.
- 9. Симмонс Л.А., Фоти Дж.Дж., Коэн С.Е., Уокер Г.К. Регулирующая сеть SOS. EcoSal Plus. 2008; 2008. Epub 2008/07/25. pmid: 25325076; Идентификатор PubMed Central PMCID: PMCPMC4196698.
- 10.Huisman O, D’Ari R. Механизм связывания индуцируемого ДНК-репликации-деления клеток в E. coli. Природа. 1981, 290 (5809): 797–9. Epub 1981/04/30. pmid: 7012641.
- 11. Huisman O, D’Ari R, Gottesman S. Контроль клеточного деления в Escherichia coli: специфическая индукция SOS-функции белка SfiA достаточно, чтобы блокировать септацию. Труды Национальной академии наук Соединенных Штатов Америки. 1984. 81 (14): 4490–4. Epub 1984/07/01. pmid: 6087326; Идентификатор PubMed Central PMCID: PMCPMC345616.
- 12. Bi E, Lutkenhaus J. Ингибиторы деления клеток SulA и MinCD предотвращают образование кольца FtsZ. Журнал бактериологии. 1993. 175 (4): 1118–25. Epub 1993/02/01. pmid: 8432706; Идентификатор PubMed Central PMCID: PMCPMC1.
- 13. Хуанг Дж., Цао С., Люткенхаус Дж. Взаимодействие между FtsZ и ингибиторами клеточного деления. Журнал бактериологии. 1996. 178 (17): 5080–5. Epub 1996/09/01. pmid: 8752322; Идентификатор PubMed Central PMCID: PMCPMC178301.
- 14. Мукерджи А., Цао С., Люткенхаус Дж.Ингибирование полимеризации FtsZ с помощью SulA, ингибитора септации у Escherichia coli. Труды Национальной академии наук Соединенных Штатов Америки. 1998. 95 (6): 2885–90. Epub 1998/04/18. pmid: 9501185; Идентификатор PubMed Central PMCID: PMCPMC19664.
- 15. Trusca D, Scott S, Thompson C, Bramhill D. Бактериальный белок контрольной точки SOS SulA ингибирует полимеризацию очищенного белка деления клеток FtsZ. Журнал бактериологии. 1998. 180 (15): 3946–53. Epub 1998/07/31. pmid: 9683493; Идентификатор PubMed Central PMCID: PMCPMC107380.
- 16. Мизусава С., Готтесман С. Распад белка в Escherichia coli: ген lon контролирует стабильность белка sulA. Труды Национальной академии наук Соединенных Штатов Америки. 1983. 80 (2): 358–62. Epub 1983/01/01. pmid: 6300834; Идентификатор PubMed Central PMCID: PMCPMC3
- .
- 17. Sonezaki S, Ishii Y, Okita K, Sugino T, Kondo A, Kato Y. Избыточное производство и очистка слитого белка SulA в Escherichia coli и его деградация протеазой Lon in vitro.Прикладная микробиология и биотехнология. 1995. 43 (2): 304–9. Epub 1995/05/01. pmid: 7612249.
- 18. Canceill D, Dervyn E, Huisman O. Протеолиз и модуляция активности ингибитора деления клеток SulA у мутантов Escherichia coli lon. Журнал бактериологии. 1990. 172 (12): 7297–300. Epub 1990/12/01. pmid: 2254289; Идентификатор PubMed Central PMCID: PMCPMC210862.
- 19. Wu WF, Zhou Y, Gottesman S. Избыточная протеолитическая активность in vivo Escherichia coli Lon и протеазы ClpYQ (HslUV).Журнал бактериологии. 1999. 181 (12): 3681–7. Epub 1999/06/15. pmid: 10368141; Идентификатор PubMed Central PMCID: PMCPMC93844.
- 20. Сон ИС, О ДжЙ, Ю СДжей, Соль Дж.Х., Чанг Ч. АТФ-зависимая деградация SulA, ингибитора клеточного деления, протеазой HslVU в Escherichia coli. Письма FEBS. 1999. 456 (1): 211–4. Epub 1999/08/19. pmid: 10452560.
- 21. Канемори М., Янаги Х., Юра Т. АТФ-зависимая протеаза HslVU / ClpQY участвует в обороте ингибитора клеточного деления SulA в Escherichia coli.Журнал бактериологии. 1999. 181 (12): 3674–80. Epub 1999/06/15. pmid: 10368140; Идентификатор PubMed Central PMCID: PMCPMC93843.
- 22. Modell JW, Hopkins AC, Laub MT. Контрольная точка повреждения ДНК в Caulobacter crescentus подавляет деление клеток за счет прямого взаимодействия с FtsW. Гены и развитие. 2011; 25 (12): 1328–43. Epub 2011/06/21. pmid: 21685367; Идентификатор PubMed Central PMCID: PMCPMC3127433.
- 23. Kawai Y, Moriya S, Ogasawara N. Идентификация белка YneA, ответственного за подавление клеточного деления во время SOS-ответа в Bacillus subtilis.Молекулярная микробиология. 2003. 47 (4): 1113–22. Epub 2003/02/13. pmid: 12581363.
- 24. Огино Х., Терамото Х., Инуи М., Юкава Х. DivS, новый SOS-индуцибельный супрессор клеточного деления в Corynebacterium glutamicum. Молекулярная микробиология. 2008. 67 (3): 597–608. Epub 2007/12/19. pmid: 18086211.
- 25. Чаухан А., Лофтон Х., Мэлони Э., Мур Дж., Фол М., Мадираджу М.В. и др. Вмешательство в деление клеток Mycobacterium tuberculosis под действием Rv2719c, гидролазы клеточной стенки. Молекулярная микробиология.2006. 62 (1): 132–47. Epub 2006/09/01. pmid: 16942606.
- 26. Modell JW, Камбара Т.К., Перчук Б.С., Лауб М.Т. SOS-независимая контрольная точка, вызванная повреждением ДНК, регулирует деление клеток Caulobacter crescentus. PLoS биология. 2014; 12 (10): e1001977. Epub 2014/10/29. pmid: 25350732; Идентификатор PubMed Central PMCID: PMCPMC4211646.
- 27.
Мо АХ, Буркхолдер ВФ. YneA, SOS-индуцированный ингибитор клеточного деления в Bacillus subtilis, регулируется посттрансляционно и требует для активности трансмембранной области.Журнал бактериологии. 2010. 192 (12): 3159–73. Epub 2010/04/20. pmid: 20400548; Идентификатор PubMed Central PMCID: PMCPMC2
5. 2; PubMed Central PMCID: PMCPMC208043.- 28. Бирн Р. Т., Чен Ш., Вуд Э. А., Кэбот Э. Л., Кокс М. М.. Гены и пути развития Escherichia coli, участвующие в выживании при экстремальном воздействии ионизирующего излучения. Журнал бактериологии. 2014. 196 (20): 3534–45. Epub 2014/07/23. pmid: 25049088; Идентификатор PubMed Central PMCID: PMCPMC4187691.
- 29. Айер В.Н., Шибальский В. Молекулярный механизм действия митомицина: связывание комплементарных цепей ДНК.Труды Национальной академии наук Соединенных Штатов Америки. 1963; 50: 355–62. Epub 1963/08/01. pmid: 14060656; Идентификатор PubMed Central PMCID: PMCPmc221180.
- 30. Нолл Д.М., Мейсон ТМ, Миллер П.С. Формирование и восстановление межцепочечных сшивок в ДНК. Химические обзоры. 2006. 106 (2): 277–301. Epub 2006/02/09. pmid: 16464006; PubMed Central PMCID: PMCPmc2505341.
- 31. Седжвик Б. Ремонт повреждений ДНК-метилированием. Обзор природы Молекулярно-клеточная биология.2004. 5 (2): 148–57. Epub 2004/03/26. pmid: 15040447.
- 32. Рейтер Х., Милевский М., Келли П. Механизм действия флеомицина на Bacillus subtilis. Журнал бактериологии. 1972; 111 (2): 586–92. Epub 1972/08/01. pmid: 4626504; Идентификатор PubMed Central PMCID: PMCPMC251321.
- 33. Кросс Дж., Хеннер В. Д., Хехт С. М., Хазелтин В. А.. Специфичность расщепления дезоксирибонуклеиновой кислоты блеомицином, флеомицином и таллисомицином. Биохимия. 1982. 21 (18): 4310–8. Epub 1982/08/31. pmid: 6181807.
- 34. Робинсон Д.Г., Чен В., Стори Д.Д., Грешем Д. Дизайн и анализ экспериментов Bar-seq. G3 (Бетесда, Мэриленд). 2014; 4 (1): 11–8. Epub 2013/11/07. pmid: 241; Идентификатор PubMed Central PMCID: PMCPMC3887526.
- 35. ван Опийнен Т., Боди К.Л., Камилли А. Tn-seq: высокопроизводительное параллельное секвенирование для изучения пригодности и генетического взаимодействия у микроорганизмов. Природные методы. 2009. 6 (10): 767–72. Epub 2009/09/22. pmid: 19767758; PubMed Central PMCID: PMCPmc2957483.
- 36.ван Опийнен Т., Камилли А. Секвенирование вставки транспозонов: новый инструмент для анализа микроорганизмов на системном уровне. Обзоры природы Микробиология. 2013. 11 (7): 435–42. Epub 2013/05/29. pmid: 23712350; PubMed Central PMCID: PMCPmc3842022.
- 37. Бенджамини Ю., Хохберг Ю. Контроль уровня ложных открытий: практичный и мощный подход к множественному тестированию. J R Stat Soc Ser B-Methodol. 1995. 57 (1): 289–300. WOS: A1995QE45300017.
- 38. Ленхарт Дж. С., Шредер Дж. В., Уолш Б. В., Симмонс Л. А..Ремонт ДНК и поддержание генома у Bacillus subtilis. Обзоры микробиологии и молекулярной биологии: MMBR. 2012. 76 (3): 530–64. Epub 2012/08/31. pmid: 22
9; PubMed Central PMCID: PMCPmc3429619.- 39. Санкар А. Эксцизионная репарация ДНК. Ежегодный обзор биохимии. 1996; 65: 43–81. Epub 1996/01/01. pmid: 8811174.
- 40. Фридман Б.М., Ясбин РЭ. Генетика и специфичность системы конститутивной эксцизионной репарации Bacillus subtilis. Молекулярная и общая генетика: MGG.1983; 190 (3): 481–6. Epub 1983/01/01. pmid: 6410154.
- 41. Уолш Б. В., Больц С. А., Вессель С. Р., Шредер Дж. В., Кек Дж. Л., Симмонс Л. А.. Хеликаза RecD2 ограничивает стресс репликационной вилки у Bacillus subtilis. Журнал бактериологии. 2014. 196 (7): 1359–68. Epub 2014/01/21. pmid: 24443534; Идентификатор PubMed Central PMCID: PMCPmc39
.- 42. Au N, Kuester-Schoeck E, Mandava V, Bothwell LE, Canny SP, Chachu K и др. Генетический состав SOS-системы Bacillus subtilis. Журнал бактериологии.2005. 187 (22): 7655–66. WOS: 000233400200013. pmid: 16267290
- 43. Горанов AI, Kuester-Schoeck E, Wang JD, Grossman AD. Характеристика глобальных транскрипционных ответов на различные типы повреждений ДНК и нарушение репликации у Bacillus subtilis. Журнал бактериологии. 2006. 188 (15): 5595–605. WOS: 0002300029. pmid: 16855250
- 44. Wu LJ, Errington J. Координация клеточного деления и сегрегации хромосом с помощью белка окклюзии нуклеоидов в Bacillus subtilis.Клетка. 2004. 117 (7): 915–25. Epub 2004/06/24. pmid: 15210112.
- 45. Горанов AI, Katz L, Breier AM, Burge CB, Grossman AD. Транскрипционный ответ на статус репликации, опосредованный консервативным бактериальным белком репликации DnaA. Труды Национальной академии наук Соединенных Штатов Америки. 2005. 102 (36): 12932–7. Epub 2005/08/27. pmid: 16120674; Идентификатор PubMed Central PMCID: PMCPMC1200305.
- 46. Брамкамп М., Уэстон Л., Дэниел Р.А., Эррингтон Дж.Регулируемый внутримембранный протеолиз белка FtsL и контроль деления клеток у Bacillus subtilis. Молекулярная микробиология. 2006; 62 (2): 580–91. Epub 2006/10/06. pmid: 17020588.
- 47. Каримова Г., Пиду Дж., Ульманн А., Ладант Д. Бактериальная двугибридная система, основанная на восстановленном пути передачи сигнала. Труды Национальной академии наук Соединенных Штатов Америки. 1998. 95 (10): 5752–6. Epub 1998/05/20. pmid: 9576956; Идентификатор PubMed Central PMCID: PMCPMC20451.
- 48. Каримова Г., Голиар Э., Дави М., Уеллетт С.П., Ладант Д. Взаимодействие белков и белков: бактериальный двугибридный вид. Методы молекулярной биологии (Клифтон, Нью-Джерси). 2017; 1615: 159–76. Epub 2017/07/02. pmid: 28667611.
- 49. Каримова Г., Даутин Н., Ладант Д. Сеть взаимодействия между мембранными белками Escherichia coli, участвующими в делении клеток, по данным бактериального двухгибридного анализа. Журнал бактериологии. 2005. 187 (7): 2233–43. Epub 2005/03/19. pmid: 15774864; Идентификатор PubMed Central PMCID: PMCPMC1065216.
- 50. Биби К.Д., Шин Дж., Пенг Дж., Чаудхури С., Кхера Дж., Пей Д. Распознавание субстрата через домен PDZ в хвостовой специфической протеазе. Биохимия. 2000. 39 (11): 3149–55. Epub 2000/03/15. pmid: 10715137.
- 51. Зильбер К.Р., Кейлер К.С., Зауэр РТ. Tsp: специфическая для хвоста протеаза, которая избирательно расщепляет белки с неполярными С-концами. Труды Национальной академии наук Соединенных Штатов Америки. 1992. 89 (1): 295–9. Epub 1992/01/01. pmid: 1729701; Идентификатор PubMed Central PMCID: PMCPMC48223.
- 52. Уолш Н.П., Альба Б.М., Бозе Б., Гросс Калифорния, Зауэр РТ. Сигналы пептида OMP инициируют стрессовую реакцию оболочки путем активации протеазы DegS посредством снятия ингибирования, опосредованного его доменом PDZ. Клетка. 2003. 113 (1): 61–71. Epub 2003/04/08. pmid: 12679035.
- 53. Сон Дж., Грант Р.А., Зауэр RT. Аллостерическая активация DegS, протеазы PDZ, сенсора стресса. Клетка. 2007. 131 (3): 572–83. Epub 2007/11/06. pmid: 17981123.
- 54. Сон Дж., Зауэр RT. Пептиды OMP модулируют активность протеазы DegS путем дифференциального связывания с активными и неактивными конформациями.Mol Cell. 2009. 33 (1): 64–74. Epub 2009/01/20. pmid: 1
28; Идентификатор PubMed Central PMCID: PMCPMC2742306. - 55. Шалтиэль И.А., Креннинг Л., Бруинсма В., Медема Р.Х. То же самое, только другое — контрольные точки повреждения ДНК и их изменение на протяжении клеточного цикла. Журнал клеточной науки. 2015; 128 (4): 607–20. Epub 2015/01/23. pmid: 25609713.
- 56. Ван Х, Чжан Х, Тэн Л., Легерски Р.Дж. Восстановление контрольной точки повреждения ДНК и развитие рака. Экспериментальные исследования клеток. 2015; 334 (2): 350–8.Epub 2015/04/07. pmid: 25842165.
- 57. Бернар Р., Маркиз К.А., Руднер Д.З. Окклюзия нуклеоидов предотвращает деление клеток во время остановки репликационной вилки у Bacillus subtilis. Молекулярная микробиология. 2010. 78 (4): 866–82. Epub 2010/09/03. pmid: 20807205; Идентификатор PubMed Central PMCID: PMCPMC2978284.
- 58. Дэниел Р.А., Гарри Э.Дж., Катис В.Л., Уэйк Р.Г., Эррингтон Дж. Характеристика основного гена деления клеток ftsL (yIID) Bacillus subtilis и его роли в сборке аппарата деления.Молекулярная микробиология. 1998. 29 (2): 593–604. Epub 1998/08/28. pmid: 9720875.
- 59. Buchholz M, Nahrstedt H, Pillukat MH, Deppe V, Meinhardt F. нестабильность мРНК yneA участвует во временном ингибировании деления клеток во время SOS-ответа Bacillus megaterium. Микробиология (Рединг, Англия). 2013; 159 (Pt 8): 1564–74. Epub 2013/06/04. pmid: 23728628.
- 60. Янгман П., Перкинс Дж. Б., Лосик Р. Создание сайта клонирования около одного конца TN917, в который может быть вставлена чужеродная ДНК, не влияя на транспозицию в Bacillus subtilis или экспрессию гена ERM транспозона-борна.Плазмида. 1984; 12 (1): 1–9. WOS: A1984Th51600001. pmid: 60
- 61. Джонсон С.М., Гроссман А.Д. Идентификация генов-хозяев, влияющих на приобретение интегративного и конъюгативного элемента в Bacillus subtilis. Молекулярная микробиология. 2014. 93 (6): 1284–301. Epub 2014/07/30. pmid: 25069588; Идентификатор PubMed Central PMCID: PMCPmc4160349.
- 62. Burby PE, Simmons LA. MutS2 способствует гомологичной рекомбинации в Bacillus subtilis. Журнал бактериологии. 2017; 199 (2). Epub 2016/11/02.pmid: 27799325; Идентификатор PubMed Central PMCID: PMCPMC5198493.
- 63. Мэтьюз Л.А., Симмонс Л.А. Взаимодействия скрытых адаптерных белков регулируют инициацию репликации ДНК. bioRxiv. 2018.
- 64. Кидане Д., Санчес Х., Алонсо Дж. К., Грауманн ПЛ. Визуализация репарации двухцепочечных разрывов ДНК у живых бактерий показывает динамическое рекрутирование белков RecF, RecO и RecN Bacillus subtilis в отдельные участки нуклеоидов. Молекулярная микробиология. 2004. 52 (6): 1627–39. Epub 2004/06/10.pmid: 15186413.
- 65. Санчес Х., Кидане Д., Кастильо Козар М., Грауманн П.Л., Алонсо Дж. Привлечение Bacillus subtilis RecN к двухцепочечным разрывам ДНК в отсутствие процессинга концов ДНК. Журнал бактериологии. 2006. 188 (2): 353–60. Epub 2005/12/31. pmid: 16385024; Идентификатор PubMed Central PMCID: PMCPMC1347269.
- 66. Санчес Х., Кидане Д., Рид П., Кертис Ф.А., Козар М.С., Грауманн П.Л. и др. Транслоказа миграции ветвей RuvAB и резольваза соединений Холлидея RecU необходимы для репарации двухцепочечных разрывов ДНК в Bacillus subtilis.Генетика. 2005. 171 (3): 873–83. Epub 2005/07/16. pmid: 16020779; Идентификатор PubMed Central PMCID: PMCPMC1456856.
- 67. Алонсо Дж. К., Людер Дж., Портной Р. Х. Характеристика рекомбинационных путей Bacillus subtilis. Журнал бактериологии. 1991. 173 (13): 3977–80. Epub 1991/07/01. pmid: 1
- 68. Mascarenhas J, Sanchez H, Tadesse S, Kidane D, Krisnamurthy M, Alonso JC и др. Белок SbcC Bacillus subtilis играет важную роль в репарации межцепочечных сшивок ДНК.BMC Mol Biol. 2006; 7:15. WOS: 000239771800001.
- 69. Санчес Х, Карраско Б., Козар М.С., Алонсо Дж. Транслоказа миграции ветвей RecG Bacillus subtilis необходима для репарации ДНК и хромосомной сегрегации. Молекулярная микробиология. 2007. 65 (4): 920–35. Epub 2007/07/21. pmid: 17640277.
- 70. Карденас П.П., Карраско Б., Дефеу Суфо С., Сезар С.Е., герр К., Кауфенштейн М. и др. RecX облегчает гомологичную рекомбинацию, модулируя активность RecA. PLoS Genet. 2012; 8 (12): e1003126.Epub 2013/01/04. pmid: 23284295; Идентификатор PubMed Central PMCID: PMCPMC3527212.
химер Drosophila melanogaster, полученных путем инъекции гаплоидных ядер
Bridges, C. B. Proc. натн. Акад. Sci. США 11 , 706–710 (1925).
ADS CAS Статья Google Scholar
Бриджес, К. Б. Science 72 , 405–406 (1930).
Google Scholar
Waletzky, E. Drosoph. Инф. Серв. 8 , 82 (1937).
Google Scholar
Sturtevant, A.H. Z. wiss. Zool. 125 , 323–356 (1929).
Google Scholar
Hinton, C. W. & McEarchen, W. Drosoph. Инф. Серв. 37 , 90 (1963).
Google Scholar
Джек, Дж. У. и Джадд, Б. Х. Proc. натн. Акад. Sci. США 76 , 1368–1372 (1979).
ADS CAS Статья Google Scholar
Ганс, М., Аудит, К. и Массон, М. Генетика 81 , 683–704 (1975).
CAS PubMed PubMed Central Google Scholar
Залокар, М., Audit, C. & Erk, I. Devl Biol. 47 , 419–432 (1975).
CAS Статья Google Scholar
Debec, A. 6th Eur. Дрозоф. Res. Конф. (тез.) (1979).
Заколар М., Эрк И. и Сантамария П. Cell 19 , 133–141 (1980).
Артикул Google Scholar
Залокар, М.И Erk, I. J. Microsc. Биол. Клетка. 25 , 97–106 (1976).
Google Scholar
Залокар, М. Прок. натн. Акад. Sci. США 68 , 1539–1541 (1971).
ADS CAS Статья Google Scholar
Сантамария, П. Арка Вильгельма Ру. Entw. Мех. Орг. 178 , 89–98 (1975).
Артикул Google Scholar
Сталкер, Х.D. Genetics 39 , 4–34 (1954).
CAS PubMed PubMed Central Google Scholar
Доан, W. Science 132 , 677–678 (1960).
ADS CAS Статья Google Scholar
Carson, H. L. Proc. натн. Акад. Sci. США 70 , 1772–1774 (1973).
ADS CAS Статья Google Scholar
Карсон, Х.L. Univ. Tex. Publs 6205 , 55–62 (1962).
Google Scholar
Subtenly, S. S. & McLoughlin, C. Anat. Рек. 125 , 623 (1956).
Google Scholar
Франкхаузер, Г. в Анализ развития (ред. Виллиер, Б. Х., Вайс, П. А. и Амбургер, Х.) 126–150 (Хафнер, Нью-Йорк, 1955).
Google Scholar
Held, L.I. Арка Вильгельма Ру. Entw Mech. Орг. 187 , 105–127 (1979).
Артикул Google Scholar
Бриджес, C. B. in Sex and Internal Secretions , 15–63 (Williams & Wilkins, Baltimore, 1932).
Google Scholar
Morata, G.