Сердечная мышечная ткань
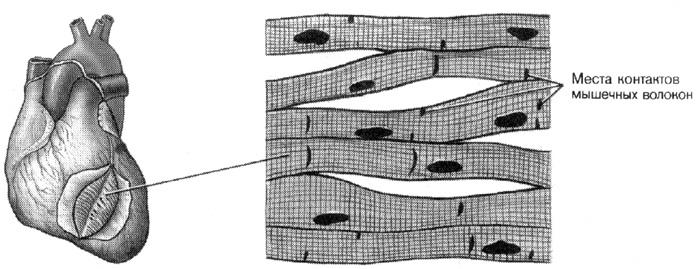
Она образует мышечную стенку сердца – миокард. Ее морфо-функциональная единица – отдельная клетка – кардиомиоцит. Клетки соединены друг с другом особыми структурами – вставочными дисками, и в результате образуется трехмерная сеть из клеточных тяжей.
Кардиомиоциты – вытянутые клетки с несколькими ответвлениями. Их ядра (1 или 2) лежат центрально.
В составе миокарда выделяют несколько видов кардиомиоцитов:
А) сократительные или рабочие
Б) проводящие
В) секреторные
Рабочие кардиомиоциты составляют основную массу миокарда и обеспечивают сокращение. Их организация сходна с мышечными волокнами, но имеет ряд отличий.
Саркотубулярная сеть развита слабее. Т-трубочки более широкие, лежат реже и каждая контактирует только с одной цистерной ЭПС (диада).
Концы соседних клеток или их ответвления соединяются вставочными дисками. Диск имеет ступенчатую форму. Поперечные участки образованы десмосомами и придают соединению механическую прочность. Продольные участки содержат множество щелевых контактов – нексусов, которых особенно много в предсердиях. Благодаря ионным каналам нексусов возбуждение быстро распространяется вдоль всей мышцы.
Миокард обильно кровоснабжается. Все промежутки между кардиомиоцитами заполнены рыхлой соединительной тканью, в которой ветвятся капилляры. Здесь же заканчиваются ветвления нервных волокон вегетативной нервной системы. На сократительную активность кардиомиоцитов нервная система оказывает лишь регуляторной влияние. Вегетативная система лишь увеличивает (симпатический отдел) или уменьшает (парасимпатический отдел) частоту и силу сердечных сокращений.
Ритмичная генерация импульсов, которые заставляют сердце постоянно сокращаться, обеспечивается специальными клетками самого миокарда. Совокупность этих клеток называется проводящей системой сердца, а способность сердца сокращаться независимо от нервных стимулов – автоматией сердца.
Проводящая система включает специализированные кардиомиоциты, называемые также атипичными. К ним относят:
Главные водители ритма – это кардиомиоциты синусно-предсердного узла. Каждую минуту они генерируют 60-90 импульсов. Водители ритма второго порядка образуют предсердно-желудочковый узел. Они генерируют импульсы с частотой 40 импульсов в минуту, и в норме их активность подавляется главными пейсмекерами. Пейсмекерные кардиомиоциты – мелкие светлые клетки с крупным ядром. Их сократительный аппарат развит слабо.
проводящие кардиомиоциты обеспечивают быструю передачу возбуждения от водителей ритма к рабочим кардиомиоцитам. Эти клетки объединены в длинные тяжи, формирующие пучок Гиса и волокна Пуркинье. Волокна Пуркинье содержат самые крупные кардиомиоциты, которые могут контактировать сразу с несколькими рабочими клетками.
Секреторные кардиомиоциты. В предсердиях встречаются отросчатые клетки, в которых хорошо развита грЭПС, комплекс Гольджи и содержатся секреторные гранулы. Миофибриллы развиты очень слабо, поскольку основной функцией является выработка гормона (натрийуретический фактор), регулирующего артериальное давление.
Сердечная мышечная ткань Википедия

 повреждение мышечной ткани миокарда
повреждение мышечной ткани миокардаМиока́рд (лат. myocardium от др.-греч. μῦς «мышца» + καρδία «сердце») — название мышечного среднего слоя сердца, составляющего основную часть его массы, требует постоянного кровоснабжения и снабжения кислородом
Строение[ | ]
Миокард образован сердечной исчерченной поперечнополосатой мышечной тканью, представляющей собой плотное соединение мышечных клеток — кардиомиоцитов, образующих основную часть миокарда. Отличается от других типов мышечной ткани (скелетная мускулатура, гладкая мускулатура) особым гистологическим строением, облегчающим распространение потенциала действия между кардиомиоцитами. Элементарной сократительной единицей кардиомиоцита является саркомер — участок миофибриллы между двумя так называемыми линиями Z. Длина саркомера равна 1,6—2,2 мкм в зависимости от степени сокращения. В саркомере чередуются светлые и тёмные полосы, отчего миофибрилла при световой микроскопии выглядит поперечно исчерченной. В центре находится тёмная полоса постоянной длины (1,5 мкм) — диск A, его ограничивают два более светлых диска I переменной длины. Саркомер миокарда, как и скелетной мышцы, состоит из переплетённых нитей (миофиламентов) двух типов. Толстые нити есть только в диске A. Они состоят из белка миозина, имеют сигарообразную форму, диаметр 10 нм и длину 1,5—1,6 мкм. Тонкие нити включают прежде всего актин и идут от линии Z через диск I в диск A. Их диаметр составляет 5 нм, длина — 1 мкм. Толстые и тонкие нити накладываются друг на друга только в диске A; диск I содержит лишь тонкие нити. При электронной микроскопии между толстыми и тонкими нитями видны поперечные мостики.
Особенности[ | ]
Главная особенность миокарда — создание ритмических движений сердца. Функциональной особенностью миокарда являются ритмичные автоматические сокращения, чередующиеся с расслаблениями, совершаются непрерывно в течение всей жизни организма. Последовательное сокращение и расслабление различных отделов сердца связано с его строением и наличием проводящей системы сердца, по которой распространяется импульс. Миокард предсердий и желудочков разобщён фиброзной перегородкой, что позволяет им сокращаться независимо друг от друга, так как возбуждение не может распространяться по фиброзной ткани. Возбуждение от предсердий к желудочкам проводится только через атриовентрикулярный пучок, отходящий от атриовентрикулярного узла [1].
Примечания[ | ]
- ↑ Гайтон А. К., Холл Д. Э. Медицинская физиология = Textbook of Medical Physiology / под ред. В.И. Кобрина. — М.: Логосфера, 2008. — С. 112. — 1296 с. — ISBN 978-5-98657-013-6.
Классификация сократимых тканей.
Государственное бюджетное общеобразовательное учреждение
Высшего профессионального образования
Тюменская государственная медицинская академия
Министерства здравоохранения и социального развития
Российской Федерации
Кафедра гистологии c эмбриологией ЗДН проф. Дунаева П.В.
РЕФЕРАТ
На тему: «Сократимые ткани мезодермального, мезенхимного, эктодермального и нейрального генеза».
Выполнила: студентка 135 группы
Вывчий А. Н.
Проверил: доцент кафедры
Истомина О. Ф.
Тюмень 2013 г.
Содержание:
Введение.
Основная часть
Классификация сократимых тканей;
Поперечно-полосатая сердечная мышечная ткань;
Поперечно-полосатая мышечная ткань скелетной мускулатуры;
Гладкая мышечная ткань;
Мышечная ткань мезенхимного происхождения;
Мышечная ткань эпидермального происхождения;
Мышечная ткань нейрального происхождения;
Целомические поперечнополосатые мышечные ткани;
Миотомные поперечнополосатые мышечные ткани;
Заключение.
Список используемой литературы.
Введение.
Клетки, из которых состоит организм человека, не одинаковы. Все они специализированы для выполнения определенных функций. Эта специализация позволяет клеткам функционировать более эффективно, но увеличивает зависимость одних частей тела от других: повреждение или разрушение одной части может привести к гибели всего организма. Вместе с тем, преимущества специализации с избытком компенсируют ее отрицательные стороны. Специализация клеток происходит уже в эмбриональном периоде развития организма и этот процесс называют дифференциацией клеток.
Группы специализированных клеток образуют ткани. Совокупность клеток и межклеточного вещества, сходных по происхождению, строению и выполняемым функциям, называют тканью. В организме человека выделяют четыре основные группы тканей: эпителиальную, соединительную, мышечную и нервную. Наука, изучающая ткани организма, называется гистологией.
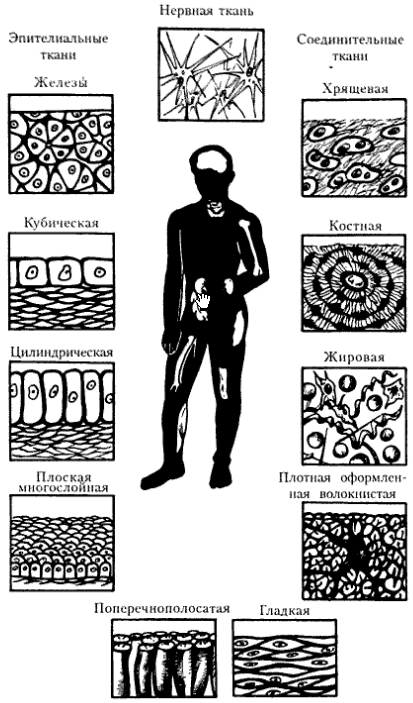
Основная часть.
Мышечными тканями (textus muscularis) называют ткани, различные по строению и происхождению, но сходные по способности к выраженным сокращениям. Они обеспечивают перемещения в пространстве организма в целом, его частей и движение органов внутри организма (сердце, язык, кишечник и др.).
Свойством изменения формы обладают клетки многих тканей, но в мышечных тканях эта способность становится главной функцией.
Основные морфологические признаки элементов мышечных тканей: удлиненная форма, наличие продольно расположенных миофибрилл и миофиламентов — специальных органелл, обеспечивающих сократимость, расположение митохондрий рядом с сократительными элементами, наличие включений гликогена, липидов и миоглобина. Специальные сократительные органеллы — миофиламенты или миофибриллы обеспечивают сокращение, которое возникает при взаимодействии в них двух основных фибриллярных белков — актина и миозина — при обязательном участии ионов кальция. Митохондрии обеспечивают эти процессы энергией. Запас источников энергии образуют гликоген и липиды. Миоглобин — белок, обеспечивающий связывание кислорода и создание его запаса на момент сокращения мышцы, когда сдавливаются кровеносные сосуды (поступление кислорода при этом резко падает).
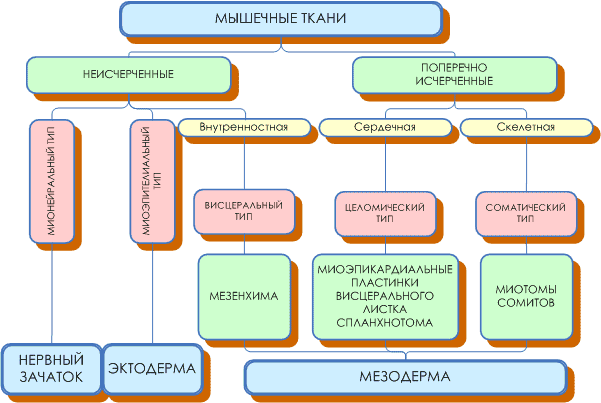
Классификация мышечных тканей:
Морфологическая
Гладкая мышечная ткань внутренних органов
Поперечно-полосатая мышечная ткань скелетной мускулатуры
Поперечно-полосатая мышечная ткань сердца
Гистогенетическая (по Н.Г. Хлопину)
Мезенхимные мышечные ткани
Эпидермальные мышечные ткани
Нейральные мышечные ткани
Мезодермальные мышечные ткани
Поперечно-полосатая сердечная мышечная ткань
Сердечная мышечная ткань, которая по строению и функции отличается от скелетных мышц, состоит из кардиомиоцитов, образующих соединяющиеся друг с другом комплексы. По своему строению сердечная мышечная ткань похожа на скелетную (поперечнополосатая исчерченность), однако сокращения сердечной мышцы не подконтрольны сознанию человека, она иннервируется вегетативной нервной системой.
Строение миофибрилл аналогично таковому скелетных мышц. Однако, в отличие от последних, между мио-фибриллами кардиомиоцитов нет столь четких границ. По периферии клетки и между митохондриями находится множество частичек гликогена и элементов гладкого эндоплазматического ретикулума. В кардиомиоцитах имеется очень большое количество крупных митохондрий с хорошо развитыми кристами, которые располагаются группами между миофибриллами. На уровне Z-линий плазмалемма кардиомиоцитов также формирует Т-трубочки, вблизи которых сосредоточены скопления цистерн гладкого эндоплазматического ретикулума. Однако триады выражены менее четко, чем в скелетных мышцах. Кардиомиоциты соединены между собой вставочными дисками . На участках вставочного диска, лежащих параллельно продольной оси кардиомиоцита , находятся лентовидные десмосомы (пояски сцепления, к ним, возможно, прикрепляются актиновые филаменты) и щелевидные контакты , не связанные с миофиламентами. Через нексусы (щелевидные контакты) осуществляются передача нервного возбуждения и обмен ионами между клетками.
Поперечно-полосатая мышечная ткань скелетной мускулатуры
Скелетная (поперечно-полосатая) мышечная ткань — упругая, эластичная ткань, способная сокращаться под влиянием нервных импульсов: один из типов мышечной ткани. Образует скелетную мускулатуру человека и животных, предназначенную для выполнения различных действий: движения тела, сокращения голосовых связок, дыхания. Мышцы состоят на 70-75 % из воды.
Источником развития скелетной мускулатуры являются клетки миотомов — миобласты. Часть из них дифференцируется в местах образования так называемых аутохтонных мышц. Прочие же мигрируют из миотомов в мезенхиму; при этом они уже детерминированы, хотя внешне не отличаются от других клеток мезенхимы. Их дифференцировка продолжается в местах закладки других мышц тела. В ходе дифференцировки возникает 2 клеточные линии. Клетки первой сливаются, образуя симпласты — мышечные трубки (миотубы). Клетки второй группы остаются самостоятельными и дифференцируются в миосателлиты (миосателлитоциты).
В первой группе происходит дифференцировка специфических органелл миофибрилл, постепенно они занимают большую часть просвета миотубы, оттесняя ядра клеток к периферии.
Клетки второй группы остаются самостоятельными и располагаются на поверхности мышечных трубок.
Структурной единицей мышечной ткани является мышечное волокно. Оно состоит из миосимпласта и миосателлитоцитов (клеток-спутниц), покрытых общей базальной мембраной.
Длина мышечного волокна может достигать нескольких сантиметров при толщине в 50-100 микрометров.

Данный вид мышечной ткани обеспечивает возможность выполнения произвольных движений. Сокращающаяся мышца воздействует на кости или кожу, к которым она прикрепляется. При этом один из пунктов прикрепления остаётся неподвижным — так называемая точка фиксации, которая в большинстве случаев рассматривается в качестве начального участка мышцы. Перемещающийся фрагмент мышцы называют подвижной точкой, которая является местом её прикрепления.
Гладкие мышцы — Википедия
Материал из Википедии — свободной энциклопедии
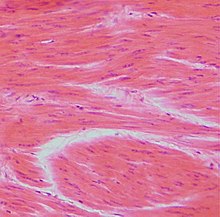 Гладко-мышечная ткань, гематоксилин-эозин.
Гладко-мышечная ткань, гематоксилин-эозин.Гладкие мышцы — сократимая ткань, в отличие от поперечнополосатых мышц не имеющая поперечной исчерченности.
Гладкие мышцы у беспозвоночных и позвоночных[править | править код]
У некоторых беспозвоночных гладкие мышцы образуют всю мускулатуру тела. У позвоночных они входят в состав оболочек внутренних органов: кишечника, кровеносных сосудов, дыхательных путей, выделительных и половых органов, а также многих желёз. Клетки гладких мышц у беспозвоночных разнообразны по форме и строению; у позвоночных в большинстве случаев веретенообразные, сильно вытянутые, с палочковидным ядром, длиной 50—250 мкм, в матке беременных животных — до 500 мкм; окружены волокнами соединительной ткани, образующими плотный футляр.
Сократимый материал — протофибриллы — обычно располагается в цитоплазме изолированно; только у некоторых животных они собраны в пучки — миофибриллы. В гладких мышцах найдены все три вида сократимого белка — актин, миозин и тропомиозин. Преимущественно встречаются протофибриллы одного типа (диаметром около 100 мкм).
Клеточных органоидов (митохондрии, комплекс Гольджи, элементы эндоплазматического ретикулума) в гладких мышцах меньше, чем в поперечнополосатой мускулатуре. Они располагаются преимущественно на полюсах ядра в цитоплазме, лишённой сократимых элементов. Клеточная мембрана часто образует карманы в виде пиноцитозных пузырьков, что указывает на резорбцию и всасывание веществ поверхностью клетки.
Установлено, что гладкие мышцы — группа различных по происхождению тканей, объединяемых единым функциональным признаком — способностью к сокращению. Так, у беспозвоночных гладкие мышцы развиваются из мезодермальных листков и целомического эпителия. У позвоночных гладкие мышцы слюнных, потовых и молочных желёз происходят из эктодермы, гладкие мышцы внутренних органов — из мезенхимы и т. д. Соседние клетки гладких мышц контактируют друг с другом отростками так, что мембраны двух клеток соприкасаются. В мышцах кишечника мыши зоны контакта занимают 5 % поверхности клеточной мембраны. Здесь, вероятно, происходит передача возбуждения от одной клетки к другой (см. Синапсы).
В отличие от поперечнополосатых мышц, для гладких мышц характерно медленное сокращение, способность долго находиться в состоянии сокращения, затрачивая сравнительно мало энергии и не подвергаясь утомлению. Двигательная иннервация гладких мышц осуществляется отростками клеток вегетативной нервной системы, чувствительная — отростками клеток спинальных ганглиев. Не каждая клетка гладких мышц имеет специализированное нервное окончание.
Мышечное сокращение — Википедия
Мы́шечное сокраще́ние — реакция мышечных клеток на воздействие нейромедиатора, реже гормона, проявляющаяся в уменьшении длины клетки. Это жизненно важная функция организма, связанная с оборонительными, дыхательными, пищевыми, половыми, выделительными и другими физиологическими процессами.
Все виды произвольных движений — ходьба, мимика, движения глазных яблок, глотание, дыхание и т. п. осуществляются за счёт скелетных мышц. Непроизвольные движения (кроме сокращения сердца) — перистальтика желудка и кишечника, изменение тонуса кровеносных сосудов, поддержание тонуса мочевого пузыря — обусловлены сокращением гладкой мускулатуры. Работа сердца обеспечивается сокращением сердечной мускулатуры.
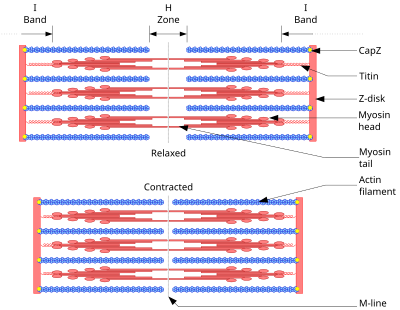 Схема, показывающая мышцы в расслабленном (выше) и сокращённом (ниже) положениях.
Схема, показывающая мышцы в расслабленном (выше) и сокращённом (ниже) положениях.Основой всех типов мышечного сокращения служит взаимодействие актина и миозина. В скелетных мышцах за сокращение отвечают миофибриллы (примерно две трети сухого веса мышц). Миофибриллы — структуры толщиной 1-2 мкм, состоящие из саркомеров — структур длиной около 2,5 мкм, состоящих из актиновых и миозиновых (тонких и толстых) филаментов и Z-дисков, соединённых с актиновыми филаментами. Сокращение происходит при увеличении концентрации в цитоплазме ионов Ca2+ в результате скольжения миозиновых филаментов относительно актиновых. Источником энергии сокращения служит АТФ. КПД мышечной клетки около 50 %, мышцы в целом не более 20%. Максимальная сила мышц не достигается в реальных условиях; не все клетки мышцы используются одновременно и сокращаются с максимальной силой, иначе при сокращении многих скелетных мышц будут повреждены сухожилия или кости (что иногда и наблюдается при сильных судорогах). КПД мышцы также зависит от внешних условий; например, на холоде он значительно снижается, так как для организма важнее сохранить температуру тела[1].
Изометрическое сокращение[править | править код]
- Основная статья: Изометрическое сокращение[en]
Изометрическое сокращение мышцы создаёт напряжение без изменения длины. Пример можно найти, когда мышцы руки и предплечья захватывают объект; суставы руки не двигаются, но мышцы генерируют достаточную силу, чтобы предотвратить падение объекта.
Изотоническое сокращение[править | править код]
- Основная статья: Изотоническое сокращение[en]
При изотоническом сокращении напряжение в мышцах остаётся постоянным, несмотря на изменение длины мышц. Это происходит, когда сила сокращения мышц соответствует общей нагрузке на мышцы.
Концентрическое сокращение[править | править код]
При концентрическом сокращении мышечное напряжение является достаточным для преодоления нагрузки, а мышца укорачивается при сокращении. Это происходит, когда сила, создаваемая мышцей, превышает нагрузку, противодействующую её сокращению.
Эксцентрическое сокращение[править | править код]
- См. также: Эксцентрическая тренировка[en]
При эксцентрическом сокращении напряжение недостаточно для преодоления внешней нагрузки на мышцы и мышечные волокна, удлиняются при их сокращении. Вместо того, чтобы тянуть сустав в направлении сокращения мышц, мышца действует так, чтобы замедлить сустав в конце движения или иным образом контролировать перемещение груза. Это может происходить невольно (например, при попытке переместить вес, слишком тяжёлый для подъёма мышцы) или добровольно (например, когда мышца «сглаживает» движение или сопротивляется гравитации, как, например, во время ходьбы вниз). В краткосрочной перспективе силовые тренировки с участием как эксцентрических, так и концентрических сокращений, по-видимому, увеличивают мышечную силу больше, чем тренировки только с концентрическими сокращениями. Однако вызванное физическими упражнениями повреждение мышц также больше при удлинении сокращений.
Эксцентрические сокращения в движении[править | править код]
Эксцентрические сокращения обычно возникают как тормозящее усилие в противоположность концентрическому сжатию, чтобы защитить суставы от повреждения. Во время практически любого обычного движения эксцентрические сокращения помогают сохранять плавность движений, но также могут замедлять быстрые движения, такие как удар или бросок. Часть тренировки для быстрых движений, таких как качки во время бейсбола, включает в себя уменьшение эксцентрического торможения, позволяющего развивать большую мощность во время движения.
Скольжение миозина относительно актина[править | править код]
Головки миозина расщепляют АТФ и за счет высвобождающейся энергии меняют конформацию, скользя по актиновым филаментам. Цикл можно разделить на 4 стадии:
- Свободная головка миозина связывается с АТФ и гидролизует его до АДФ и фосфата и остаётся связанной с ними. (Обратимый процесс — энергия, выделившаяся в результате гидролиза, запасается в изменённой конформации миозина).
- Головки слабо связываются со следующей субъединицей актина, фосфат отделяется, и это приводит к прочному связыванию головки миозина с актиновым филаментом. Эта реакция уже необратима.
- Головка претерпевает конформационное изменение, производящее подтягивание толстого филамента к Z-диску (или, что эквивалентно, свободных концов тонких филаментов друг к другу).
- Отделяется АДФ, за счёт этого головка отделяется от актинового филамента. Присоединяется новая молекула АТФ.
Далее цикл повторяется до уменьшения концентрации ионов Ca2+ или исчерпании запаса АТФ (в результате смерти клетки). Скорость скольжения миозина по актину ≈15 мкм/сек. В миозиновом филаменте много (около 500) молекул миозина и, следовательно, при сокращении цикл повторяется сотнями головок сразу, что и приводит к быстрому и сильному сокращению. Следует заметить, что миозин ведёт себя как фермент — актин-зависимая АТФаза. Так как каждое повторение цикла связано с гидролизом АТФ, а следовательно, с положительным изменением свободной энергии, то процесс однонаправленный. Миозин движется по актину только в сторону плюс-конца.
Последовательные стадииДля сокращения мышцы используется энергия гидролиза АТФ, но мышечная клетка имеет крайне эффективную систему регенерации запаса АТФ, так что в расслабленной и работающей мышце содержание АТФ примерно равно. Фермент фосфокреатинкиназа катализирует реакцию между АДФ и креатинфосфатом, продукты которой — АТФ и креатин. Креатинфосфат содержит больше запасённой энергии, чем АТФ. Благодаря этому механизму при вспышке активности в мышечной клетке падает содержание именно креатинфосфата, а количество универсального источника энергии — АТФ — не изменяется. Механизмы регенерации запаса АТФ могут различаться в зависимости от парциального давления кислорода в окружающих тканях (см. Анаэробные организмы).
В основном в регуляции мышечной активности участвуют нейроны, но есть случаи, когда сокращением гладкой мускулатуры управляют и гормоны (например, адреналин и окситоцин). Сигнал о сокращении можно разделить на несколько этапов:
От клеточной мембраны до саркоплазматического ретикулума[править | править код]
Воздействие медиатора, выделившегося из мотонейрона, вызывает потенциал действия на клеточной мембране мышечной клетки, который передаётся далее с помощью специальных впячиваний мембраны, называемых Т-трубочками, которые отходят от мембраны внутрь клетки. От Т-трубочек сигнал передаётся саркоплазматическому ретикулуму — особому компартменту из уплощенных мембранных пузырьков (эндоплазматической сети мышечной клетки), окружающих каждую миофибриллу. Этот сигнал вызывает открытие Ca2+-каналов в мембране ретикулума. Обратно ионы Ca2+ попадают в ретикулум с помощью мембранных кальциевых насосов — Ca2+-АТФазы.
От выделения ионов Ca2+ до сокращения миофибрилл[править | править код]
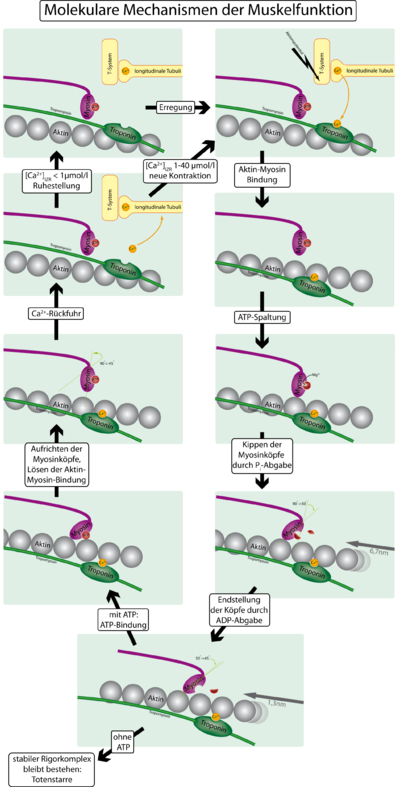 Механизм сокращения мышц с учётом тропонина и тропомиозина
Механизм сокращения мышц с учётом тропонина и тропомиозинаДля того, чтобы контролировать сокращение, к актиновому филаменту прикрепляется белок тропомиозин и комплекс из трёх белков — тропонин (субъединицы этого комплекса называются тропонинами T,I и C). Тропонин C — близкий гомолог другого белка, кальмодулина. Через каждые семь субъединиц актина расположен только один тропониновый комплекс. Связь актина с тропонином I перемещает тропомиозин в положение, мешающее связи миозина с актином. Тропонин C связывается с четырьмя ионами Ca2+ и ослабляет действие тропонина I на актин, и тропомиозин занимает положение, не препятствующее связи актина с миозином. Источником энергии для сокращения мышечных волокон служит АТФ. При связывании тропонина с ионами кальция активируются каталитические центры для расщепления АТФ на головках миозина. За счет ферментативной активности головок миозина гидролизуется АТФ, расположенный на головке миозина, что обеспечивает энергией изменение конформации головок и скольжение нитей. Освобождающиеся при гидролизе АТФ молекула АДФ и неорганический фосфат используются для последующего ресинтеза АТФ. К миозиновой головке присоединяется новая молекула АТФ. При этом происходит разъединение поперечного мостика с нитью актина. Повторное прикрепление и отсоединение мостиков продолжается до тех пор, пока концентрация кальция внутри миофибрилл не снизится до подпороговой величины. Тогда мышечные волокна начинают расслабляться.
| Белок | Доля белка % | Его мол. масса, kDa | Его функция |
|---|---|---|---|
| Миозин | 44 | 510 | Главный компонент толстых филаментов. Образует связи с актином. Движется по актину за счёт гидролиза АТФ. |
| Актин | 22 | 42 | Главный компонент тонких филаментов. Во время сокращения мышцы по нему движется миозин. |
| Титин | 9 | 2500 | Большой гибкий белок, образующий цепь для связывания миозина с Z-диском. |
| Тропонин | 5 | 78 | Комплекс из трёх белков, регулирующий сокращение при связывании с ионами Ca2+. |
| Тропомиозин | 5 | 64 | Связанный с актиновыми филаментами стержневидный белок, блокирующий движение миозина. |
| Небулин[en] | 3 | 600 | Длинный нерастяжимый белок, связанный с Z-диском и идущий параллельно актиновым филаментам. |
- Б. Альбертс, Д. Брей, Дж. Льюис, М. Рефф, К. Робертс, Дж. Уотсон, Молекулярная биология клетки — В 3-х т. — Пер. с англ. — Т.2. — М.: Мир, 1994. — 540 с.
- М. Б. Беркинблит, С. М. Глаголев, В. А. Фуралев, Общая биология — В 2-х ч. — Ч.1. — М.:МИРОС, 1999. — 224 с.: ил.