Безъядерные клетки человека
Всем известно, что человек является эукариотом. Это значит, что все его клетки имеют органеллу, в которой заключена вся генетическая информация, — ядро. Однако существуют и исключения. Есть ли в организме человека безъядерные клетки и каково их значение для жизнедеятельности?
Безъядерные клетки человека
Их нельзя сравнивать с прокариотами, обладающими типичным строением. Что же это за безъядерные клетки? Ядра нет в клетках крови — эритроцитах. Вместо данной органеллы они содержат сложный химический комплекс веществ, позволяющий им выполнять важнейшие для организма функции. Кровяные пластинки — тромбоциты и лимфоциты — также безъядерные клетки. Ядра нет и в клетках, которые называют стволовыми. Все перечисленные структуры объединяет еще один признак. Поскольку в них отсутствует ядро, они не способны к размножению. Это значит, что безъядерные клетки, примеры которых были приведены, после выполнения своей функции гибнут, а новые образуются в специализированных органах.

Эритроциты
Именно они определяют цвет нашей крови. Безъядерные клетки крови эритроциты имеют необычную форму — двояковогнутого диска, которая значительно увеличивает их поверхность при относительно малых размерах. Зато количество их просто поражает: в 1 кв. мм крови их находится до 5 млн! В среднем эритроцит живет до четырех месяцев, после чего погибает и нейтрализуется в селезенке и печени. Новые клетки формируются каждую секунду в красном костном мозге.

Функции эритроцитов
Что же вместо ядра содержат эти безъядерные клетки? Называются эти вещества гем и глобин. Первое является железосодержащим. Оно не только окрашивает кровь в красный цвет, но и образует нестойкие соединения с кислородом и углекислым газом. Глобин представляет собой вещество белковой природы. В его крупную молекулу погружен гем, содержащий заряженный ион железа. По механизму действия эти клетки можно сравнить с маршрутным такси. В легких они присоединяют кислород. С током крови он разносится ко всем клеткам и высвобождается там. При участии кислорода происходит процесс окисления органических веществ с выделением определенного количества энергии, которую человек использует для осуществления жизнедеятельности. Освободившееся место тут же занимает углекислый газ, который движется в обратном направлении — в легкие, где выдыхается. Этот процесс является необходимым условием жизни. Если кислород не поступает к клеткам, происходит их постепенное отмирание. Это может быть опасным для жизни организма в целом.
Эритроциты выполняют еще одну важную функцию. На их мембранах находится белковый маркер, который называется резус-фактором. Этот показатель, как и группа крови, очень важен во время переливания крови, при беременности, донорстве и хирургических операциях. Его обязательно устанавливают, поскольку при несовместимости может произойти так называемый резус-конфликт. Он является защитной реакцией, но может привести к отторжению плода или органов.

Нерациональное питание, вредные привычки, загрязненный воздух могут вызвать разрушение эритроцитов. Вследствие этого возникает тяжелое заболевание, которое называется анемией, или малокровием. При этом человек чувствует головокружение, слабость, одышку, шум в ушах. Кислородная недостаточность негативно сказывается на физической и умственной деятельности человека. Особенно опасна она в период беременности. Если через пуповину к плоду поступает недостаточно кислорода, это может привести к серьезным нарушениям в его развитии.
Строение тромбоцитов
Безъядерные клетки тромбоциты еще называют кровяными пластинками. В неактивном состоянии они действительно имеют плоскую форму, напоминающую линзу. А вот при повреждении сосудов они набухают, округляются, образуют непостоянные выросты наружного слоя — псевдоподии. Тромбоциты образуются в красном костном мозге и живут недолго — до 10 дней, обезвреживаясь в селезенке.

Процесс образования тромба
Матрикс кровяных пластинок содержит фермент, который называется тромбопластином. При нарушении целостности сосудов он оказывается в плазме. Под его действием белок крови протромбин переходит в свою активную форму, в свою очередь, действуя на фибриноген. В результате это вещество переходит в нерастворимое состояние. Оно превращается в белок фибрин. Его нити тесно переплетаются и образуют тромб. Защитная реакция свертывания крови предотвращает кровопотери. Однако образование тромба внутри сосуда очень опасно. Это может привести к его разрыву и даже гибели организма. Нарушение процесса свертываемости называется гемофилией. Это наследственное заболевание характеризуется недостаточным количеством тромбоцитов и приводит к излишней потере крови.

Стволовые клетки
Эти безъядерные клетки называются стволовыми не зря. Они действительно являются основой для всех других. Их еще называют «генетически чистыми». Стволовые клетки находятся во всех тканях и органах, но больше всего их содержит костный мозг. Они способствуют восстановлению целостности там, где это необходимо. Стволовые превращаются в любые другие типы клеток при их разрушении. Казалось бы, при наличии такого волшебного механизма человек должен жить вечно. Почему же этого не происходит? Все дело в том, что с возрастом интенсивность дифференциации стволовых клеток значительно уменьшается. Они уже неспособны восстановить разрушенные ткани. Но есть и еще одна опасность. Существует большая вероятность превращения стволовых клеток в раковые, что неминуемо приведет к гибели любой живой организм.

Безъядерные клетки: примеры и черты отличия
В природе безъядерные клетки встречаются достаточно часто. Например, прокариотическими являются сине-зеленые водоросли и бактерии. Но, в отличие от безъядерных клеток человека, они не гибнут после выполнения своей биологической роли. Дело в том, что прокариоты имеют генетический материал. Поэтому они способны к делению, которое происходит путем митоза. В результате образуются две генетические копии материнской клетки. Наследственная информация прокариот представлена кольцевой молекулой ДНК, которая удваивается перед делением. Этот аналог ядра еще называют нуклеоидом. У растений безъядерными являются живые клетки проводящей ткани — ситовидные трубки.
Итак, безъядерные клетки человека неспособны к делению, поэтому они существуют непродолжительный промежуток времени до выполнения своей функции. После этого происходит их разрушение и внутриклеточное переваривание. К ним относятся форменные элементы (эритроциты), кровяные пластинки (тромбоциты) и стволовые клетки.
особенности строения, примеры :: SYL.ru
Биология изучает все живое на планете Земля, начиная с глобальной экосистемы Земли — биосферы — и заканчивая самыми мельчайшими живыми частицами — клетками. Раздел биологии о клетках называется «цитология». Она изучает все живые клетки, которые бывают ядерными и безъядерными.

Значение ядра для клетки
Как видно из названия, безъядерные клетки не имеют ядра. Они характерны для прокариотов, которые сами по себе являются такими клетками. Сторонники теории эволюции считают, что эукариотические клетки произошли от прокариотических. Основным отличием эукариотов в процессе развития жизни стало именно клеточное ядро. Дело в том, что в ядрах содержится вся наследственная информация – ДНК. Потому для эукариотических клеток отсутствие ядра обычно отклонение от нормы. Однако бывают исключения.
Прокариотические организмы
Безъядерными клетками являются прокариотические организмы. Прокариоты – древнейшие существа, состоящие из одной клетки или колонии клеток, к ним относятся бактерии и археи. Их клетки называют доядерными.

Главной особенностью биологии клеток прокариотов является, как уже было упомянуто, отсутствие ядра. По этой причине их наследственная информация хранится оригинальным способом – вместо эукариотических хромосом ДНК прокариота «упакована» в нуклеоид – кольцевую область в цитоплазме. Наряду с отсутствием оформленного ядра нет мембранных органоидов – митохондрий, аппарата Гольджи, пластид, эндоплазматической сети. Вместо них необходимые функции выполняются мезосомами. Рибосомы прокариотов гораздо меньше эукариотических по размеру, а их количество меньше.
Безъядерные клетки растений
У растений есть ткани, состоящие из одних безъядерных клеток. Например, луб или флоэма. Он находится под покровной тканью и представляет собой систему из разных тканей: основной, опорной и проводящей. Основным элементом луба, относящимся к проводящей ткани, являются ситовидные трубки. Состоят они из члеников — удлинённых безъядерных клеток с тонкими клеточными стенками, главным компонентом которых являются целлюлоза и пектиновые вещества. Ядро они теряют при созревании — оно отмирает, а цитоплазма превращается в тонкий слой, размещённый у стенки клетки. Жизнь этих безъядерных клеток связана с клетками-спутниками, имеющими ядро; они тесно связаны друг с другом и фактически составляют одно целое. Членики и спутники развиваются в общей меристематической клетке.

Клетки ситовидных трубок живые, но это единственное исключение; все остальные клетки без ядра у растений являются мертвыми. У эукариотических организмов (к которым относятся и растения) безъядерные клетки способны жить очень короткое время. Клетки ситовидных трубок недолговечны, после смерти образуют поверхностный слой растения – покровную ткань (например, кору дерева).
Безъядерные клетки человека и животных
В организме человека и млекопитающих животных также есть клетки без ядра – эритроциты и тромбоциты. Рассмотрим их подробнее.
Эритроциты
Иначе их называют красными кровяными тельцами. На этапе формирования молодые эритроциты содержат ядро, а вот взрослые клетки его не имеют.
Эритроциты обеспечивают насыщение кислородом органов и тканей. С помощью содержащегося в красных кровяных клетках пигмента гемоглобина клетки связывают молекулы кислорода и переносят их от лёгких в мозг и к другим жизненно важным органам. Также они участвуют в выводе из организма продукта газообмена – углекислого газа СО2, транспортируя его.

Эритроциты человека имеют размер всего 7-10 мкм и форму двояковогнутого диска. Благодаря маленьким размерам и эластичности, красные кровяные тельца легко проходят через капилляры, которые значительно меньше них по размеру. В результате отсутствия ядра и других клеточных органелл количество гемоглобина в клетке повышено, гемоглобин заполняет весь её внутренний объём.
Выработка эритроцитов проходит в костном мозге ребёр, черепа и позвоночника. У детей задействован также костный мозг костей ног и рук. Каждую минуту формируется более 2 миллионов эритроцитов, живущих около трёх месяцев. Интересный факт – красные клетки крови составляют примерно ¼ от всех клеток человека.
Тромбоциты
Раньше их называли еще кровяными пластинками. Это мелкие безъядерные клетки крови плоской формы, размер которых не превышает 2-4 мкм. Представляют собой фрагменты цитоплазмы, которые отделились от клеток костного мозга – мегакариоцитов.

Функцией тромбоцитов является формирование сгустка крови, который «затыкает» в сосудах поврежденные места, и обеспечение нормальной свертываемости крови. Также кровяные пластинки могут выделять соединения, способствующие росту клеток (так называемые факторы роста), поэтому они важны для заживления поврежденных тканей и способствуют их регенерации. Когда тромбоциты активизируются, то есть переходят в новое состояние, они принимают форму сферы с выростами (псевдоподиями), при помощи которых сцепляются друг с другом или сосудистой стенкой, закрывая тем самым её повреждение.
Отклонение количества тромбоцитов от нормы может приводить к различным заболеваниям. Так, уменьшение количества кровяных пластинок повышает риск кровотечений, а их увеличение приводит к тромбозу сосудов, то есть появлению сгустков крови, которые в свою очередь могут стать причиной инфарктов и инсультов, эмболии лёгочной артерии и закупорке сосудов в других органах.
Образуются тромбоциты в костном мозге и селезёнке. После формирования 1/3 из них разрушается, а оставшиеся циркулируют в кровотоке чуть дольше недели.
Корнеоциты
Некоторые клетки кожи человека также не содержат ядер. Из безъядерных клеток состоят два верхних слоя эпидермиса – роговой и блестящий (цикловидный). Оба состоят из одинаковых клеток – корнеоцитов, которые представляют собой бывшие клетки нижних слоев эпидермиса – кератиноциты. Эти клетки, образовавшись на границе наружного и среднего слоев кожи (дермы и эпидермиса), поднимаются по мере «взросления» все выше, в шиповатый, а затем и в зернистый слои эпидермиса. В кераноците накапливается вырабатываемый им белок кератин — важный компонент, который отвечает за прочность и упругость нашей кожи. В итоге клетка теряет ядро и практически все органеллы, поэтому большую её часть составляет белок кератин.

Получившиеся корнеоциты имеют плоскую форму. Плотно прилегая друг к другу, они образуют роговой слой кожи, служащий барьером для микроорганизмов и многих веществ – его чешуйки выполняют защитную функцию. Переходным от зернистого к роговому служит блестящий слой, также состоящий из потерявших ядра и органеллы кератиноцитов. По сути, корнеоциты – это мертвые клетки, так как никаких активных процессов в них не происходит.
Безъядерные клетки в трансплантологии
Для клонирования клеток нужных тканей в трансплантологии используются искусственно созданные безъядерные клетки. Так как генетическую информацию у эукариотических организмов хранит именно ядро, путём манипуляций с ним можно воздействовать на свойства клетки. Как бы фантастически это ни звучало, но можно заменить ядро и таким способом получить совершенно другую клетку. Для этого ядра удаляются или разрушаются различными способами – хирургическим, с помощью ультрафиолетового излучения или центрифугирования в сочетании с воздействием цитохалазинов. В полученную безъядерную клетку пересаживают новое ядро.
До сих пор учёные не пришли к общему мнению по поводу этичности клонирования, потому оно всё ещё находится под запретом.

Таким образом, фактически живые безъядерные клетки у высших (эукариотических) организмов почти не встречаются. Исключением являются клетки крови человека – эритроциты и тромбоциты, а также клетки флоэмы у растений. В остальных случаях безъядерные клетки нельзя назвать живыми, как, например, клетки верхних слоев эпидермиса или клетки, полученные искусственным путем для клонирования тканей в трансплантологии.
Сайт учителей биологии МБОУ Лицей № 2 города Воронежа
Форменные элементы крови. Эритроциты
Эритроциты (греч. erythros — красный, cytos (kytos) — клетка), или красные кровяные клетки, — безъядерные форменные элементы крови, содержащие гемоглобин. Имеют форму двояковогнутого диска. Отличаются гибкостью и эластичностью, легко деформируются и проходят через кровеносные капилляры с диаметром меньшим, чем диаметр эритроцита. Образуются в красном костном мозге, разрушаются в печени и селезёнке. Продолжительность жизни 100 — 120 дней. В норме в 1 мкл (мм3) крови у мужчин содержится 4 — 5 млн эритроцитов, у женщин — 3,7 — 4,7 млн, у новорождённых до 6 млн.1. Дыхательная — благодаря гемоглобину переносят О2 и СО2.
2. Питательная — переносят на своей поверхности аминокислоты и доставляют их клеткам организма.
3. Буферная — поддержание с помощью гемоглобина рН крови.
4. Регуляторная
Окраска эритроцитов зависит от содержащегося в них гемоглобина. Гемоглобин способен легко соединяться с кислородом и легко отдавать его. Красные кровяные клетки переносят кислород от легких ко всем органам.
отличие от других классов позвоночных животных у млекопитающих (и человека в том числе) эритроциты не имеют ядра. Оно выталкивается по мере созревания клетки, и эритроцит приобретает двояковогнутую форму, что увеличивает поверхность соприкосновения эритроцита с воздухом легочных пузырьков и увеличивает его полезный объем (ядро эритроцита других позвоночных гемоглобин не содержит!). Углекислый газ от тканей к легким частично переносит плазма крови, частично – гемоглобин эритроцитов.
Гемоглобин — сложное химическое соединение, которое находится в эритроцитах и состоит из двух частей: белка глобина и четырёх молекул гема. Таким образом, гемоглобин — это сложный белок — хромопротеид. Гем имеет в своём составе атом железа, способный присоединять и отдавать молекулу кислорода. При этом валентность железа не изменяется (оно остаётся двухвалентным).
В капиллярах лёгких гемоглобин присоединяет кислород и становится оксигемоглобином, который придаёт артериальной (богатой кислородом) крови ярко-алый цвет. В капиллярах тканей гемоглобин отдаёт кислород и превращается в восстановленный гемоглобин, который придаёт венозной крови тёмно-вишнёвый цвет. При соединении гемоглобина с углекислым газом (CO2) образуется карбогемоглобин.
При действии на гемоглобин сильных окислителей железо окисляется и переходит из двухвалентного в трёхвалентное, а гемоглобин превращается в метгемоглобин, придающий крови коричневую окраску. Гемоглобин очень легко соединяется с угарным газом (СО), при этом образуется карбоксигемоглобин
При разрушении эритроцитов гемоглобин выходит в плазму крови, которая окрашивается при этом в красный цвет и становится прозрачной — «лаковая кровь». Процесс разрушения эритроцитов и выхода гемоглобина носит название гемолиз. Он может произойти при уменьшении осмотического давления, что вначале приводит к набуханию, а затем разрушению эритроцитов, при воздействии химических веществ, разрушающих оболочку эритроцитов (эфир, хлороформ, алкоголь, желчные кислоты и т. д.), при сильных механических воздействиях на кровь, например при её транспортировке, при замораживании и размораживании крови, при её нагревании до 65 — 68оС, при переливании несовместимой или недоброкачественной крови, при укусах ядовитых змей, скорпионов, при иммунных реакциях.
В скелетных мышцах и миокарде (сердечной мышце) находится мышечный гемоглобин — миоглобин. Он отличается от гемоглобина крови меньшей молекулярной массой глобина (белковой части). Назначение миоглобина — снабжение кислородом работающей мышцы в момент сокращения, когда кровоток в ней уменьшен или прекращается вовсе.
< Предыдущая страница «Состав и функции крови»
Следующая страница «Лейкоциты» >
ФОРМЕННЫЕ ЭЛЕМЕНТЫ КРОВИ. ЭРИТРОЦИТЫ
ФОРМЕННЫЕ ЭЛЕМЕНТЫ КРОВИ. ЭРИТРОЦИТЫ
В висячей капле крови эритроциты находятся в таком огромном количестве, что закрывают первое время все другие элементы. Они сообщают крови её красный цвет и непрозрачность. При разрушении эритроцитов дестиллированной водой красный цвет усиливается, но кровь становится совершенно прозрачной. Разрушение эритроцитов происходит в любой жидкости, молекулярная концентрация которой отличается от сыворотки, но наибольшее разрушающее действие оказывает дестиллированная вода.
Красные кровяные тельца, при полном отсутствии, амёбовидных движений, отличаются исключительной мягкостью, гибкостью и эластичностью. Проходя через капилляры, эритроциты сильно вытягиваются в длину, но как только выходят в широкое русло, снова мгновенно превращаются в диски. Под покровным стеклом эритроциты, в первый момент, пока есть ток жидкости, сталкиваясь друг с другом, сплющиваются, изменяют форму, но как только ток прекращается, они принимают свою обычную форму.
Типичной формой эритроцитов большинства видов млекопитающих является форма равномерно закруглённых, двояковогнутых, безъядерных пластинок. При рассмотрении их в профиль они имеют форму бисквитов, а en face—вид тарелки с более тёмной периферией и светлоокрашенным центром.
Иногда эритроциты выглядят не как двояковогнутая чечевица, а как плосковогнутая (форма блюдца). Реже встречаются эритроциты, одна сторона которых вогнута, а другая, напротив, выпукла (выпукло-вогнутая). Такие эритроциты имеют вид колокола или чаши. Некоторые авторы считают1, что две последние формы являются истинными, а форма двояковогнутой чечевицы—искусственной. Однако большинство исследователейе! придерживается противоположного взгляда, считан колоколообразную форму и форму блюдца артефактами.
У верблюда, оленя п ламы эритроциты периферической крови имеют овальную форму. В костном мозгу верблюда, по данным Галузо, основная масса эритроцитов имеет круглую форму, но к моменту перехода в периферическую кровь круглоядерные эритроциты верблюда принимают эллипсовидную форму.
В крови, размазанной на предметном стекле, эритроциты часто имеют вид монетных столбиков, в которых каждый эритроцит обычно наполовину прикрывается рядом с ним лежащим. Это явление объясняется снятием заряда или уменьшением его вследствие увеличения щёлочности.
Отдельные эритроциты имеют желтовато-зеленоватый цвет, а толстый слой их кажется красноватым.
Величина эритроцитов у различных животных различна. Средний диаметр эритроцитов (в микронах) следующий:
Эритроциты состоят из нежной липоидной стромы, которая заполнена гемоглобином.
Во периферии эритроцита имеется уплотнённый слой липоидного характера, образующий подобие его оболочки. Гемоглобин, как коллоидное вещество, не может проходить через эту оболочку; она проницаема только для воды и газов. Благодаря этой полупроницаемой мембране, эритроцит удерживает свой специфический состав солей.
В свежем состоянии эритроцит представляется совершенно гомогенным и бесструктурным. Несмотря на это, издавна высказывалось предположение, что определённая структура в эритроците существует, но отдельные составные части её обладают одинаковым показателем преломления, и потому не отличимы друг от друга.
Во Брюкке, основой вещества эритроцита является сетчатый или губчатый остов—строма (ойкоид, дископлазма и т. п.), соответствующая спонгиоплазме других клеток. В петлях стромы находится гемоглобин. В пользу этой теории защитники её указывали на разнообразные сетчатые структуры, наблюдаемые при особых методах фиксации и окрашивания. Вначале они совершенно отрицали оболочку в эритроците, но затем, ввиду накопившихся в науке доказательств наличия оболочки, допустили существование на периферии эритроцита губки или сетки, спаянной с перекладинами стромы.
Шванн высказал теорию, по которой эритроциты млекопитающих сравниваются с пузырьками, имеющими мягкую, эластичную липоидную оболочку, наполненную аморфным жидким веществом—эндозомой, с растворённым в ней гемоглобином. Эта старая теория нашла защитников среди современных учёных, которые, помещая каплю костяного масла в слабый спирт одинакового с маслом удельного веса, получили модель красного кровяного тельца с липоидной оболочкой и окрашенным жидким содержимым. В качестве главного аргумента сторонники этой теории выдвигают то, что эритроциты относятся к изменениям осмотического давления как пузыри с жидким содержимым -и полупроницаемой липоидной оболочкой. Наличие оболочки у эритроцитов один из них доказал, воздействуя на эритроциты раствором таннина. Под влиянием гипотонического раствора эритроциты набухают, затем их оболочка лопается, а жидкое содержимое вытекает через разрыв, под влиянием таннина, свёртывается и остаётся приклеенным к оболочке эритроцита в виде зернистого тельца. По Заварзину, оболочку эритроцита можно нарушить микроманипулятором, после чего содержимое эритроцита изливается в окружающую среду.
Шиллинг пришёл к выводу, что, по крайней мере, молодая форма эритроцитов представляет собой клетку сложного строения. Гипотетическое строение эритроцита млекопитающих, по Шиллингу, такое:
1) протоплазм а—состоит из радиальной структуры, базофильной субстанции (в юном возрасте) и коркообраз-ной наружной оболочки;
2) архоплазма—из светлого центрального вещества (стекловидное тело) и микроцентра в соединении с прилежащим шаровидным «капсульным телом»;
3) ядро или его остатки.
К признанию сложности строения эритроцита склоняются в последнее время многие авторы.
Эритроциты постного Мозга. Клетки эритробластиче-ского ряда содержат ядро и называются э р и т р об л а с т а м и (Максимов, 1910 г.).
Эритробласты не одинаковы на полюсах своего развит тин и не резко отграничены друг от друга в ближайших звеньях. Это зависит от того, что имеются переходные формы, которые представляют собой последовательные стадии развития одной и той же клеточной формы. Можно построить непрерывный ряд клеток, от молодой клетки, бедной гемоглобином, до зрелого безъядерного эритроцита.
Большинство исследователей считает, что развитие эритроцитов идёт по двум линиям, соответствующим нормальному и патологическому эритропоэзу.
Нормобласты происходят из мезенхимальных клеток и дают начало нормальному эритропоэзу, а мегалобласты—из клеток эндотелия сосудов (мегалобластическое кроветворение, которое наблюдатся при некоторых патологических состояниях, например, при лейкемии).
По некоторым авторам, эритробласт, созревая, теряет ядро и переходит в макроцит, а затем в нормоцит; мегалобласт же с потерей ядра переходит в мегалоцит.
Мегалобласты и эритробласты—два абсолютно различных вида клеток. Эта точка зрения разделяется Горяевым, Артыновым, Ткаченко и многими другими исследователями.
Молодая форма эритроцитов различными авторами называется поразному: полихроматофильным эритробластом, мегалобластом, макронормобластом, проэритробла-стом, эритрогонием, макробластом и лимфоидным эритробластом.
В литературеописываются различные схемы эритропоэза, причём переход к зрелым формам в одних схемах двух-и трёхстепенный, в других—четырёх- и даже многостепенный.
Практически наиболее удобней является схема, приведённая Владос, которая была использована в работах Ткаченко, Смирнова, Васильева, Колесова и др. при изучении крови сельскохозяйственных животных.
По Владос, из материнской клетки образуется проэритробласт—пред стадия эритроцита. По мере созревания проэрит робласт переходит в эритробласт, а последний трансформируется в нормобласт и затем в нормоцит.
Переход из одной формы в другую нужно понимать не в смысле непосредственного превращения одной и той же клетки, а в смысле постепенного изменения структуры и характера каждого нового поколения клеток, возникающих при делении более молодых форм. С каждым делением получаются дочерние клетки меньшего размера с большим количеством протоплазмы и гемоглобина. Ядро, напротив, становится меньше, теряет способность делиться и становится постепенно пикнотичным. В дальнейшем ядро исчезает, и из нормобласта получается зрелая клетка—нормоцит.
Процесс утраты ядра нормобластами трактуется различно. Одни авторы указывают, что в костном мозгу, наряду с ядерными, имеются и безъядерные эритроциты, полагая, что у последних ядро исчезает внезапно, выталкивается или выскальзывает из клетки. При этом ядро может исчезать сразу или частями, если оно предварительно распалось. Место, где было ядро, западает; этим и объясняется образование формы эритроцитов в виде двояковогнутой чечевицы. Другие авторы полагают, что пикно-тическое ядро подвергается внутриклеточному растворению. Хроматин ядра выщелачивается, масса ядра становится меньше и меньше и, наконец, исчезает совершенно. Из шарообразного эритроцит становится плоским, с одним или двумя вдавлениями. Третьи авторы считают, что ядро сначала распадается на отдельные части, которые затем рассасываются.
В костном мозгу можно отметить как распад, так и растворение ядра. Что касается выталкивания, то оно, возможно, связано с механическим воздействием. Но если допуститВ, что имеет место выталкивание ядра или его выскальзывание, то тогда можно было бы найти очень много голых ядер, чего в действительности не наблюдается. «Кроме того, трудно представить себе, чтобы атрофированное ядро было способно к активному действию в клетке и оставалось ещё жизнеспособным после энуклеации» (Ткаченко).
Проэритробласт—самая молодая клетка костного мозга. Диаметр её у лошади, по данным Ткаченко, от 17 до 20 [I.
Проэритробласт имеет неправильную круглую форму с большим круглым ядром, расположенным или в центре клетки или эксцентрично.
Рис. 1. Кровь лошади.
Ядро проэритробласта рыхлое, имеет инжносетчатое строение, с неясно диференцированным рисунком хроматина. Хроматин ядра в виде угловатых глыбок, приблизительно одинаковой величины, образует грубую шагреневость. Во всех ядрах имеются от 2 до 4, различной величины, нуклеолей, окрашивающихся в серовато-синий цвет со слабо фиолетовым оттенком. По мере созревания проэритробласта, нуклеоли уменьшаются и исчезают.
Протоплазма проэритробласта сплошь зернистая, при окраске—интенсивно синяя, базофильная, иногда о зеленоватым оттенком. У отдельных тел ток, главным образом, созревающих, местами можно отметить серовато-желтоватое просветление. Перинуклеарная зона отсутствует. В местах более широкого пояса, в средней части протоплазмы, отмечаются светлые, бледноокрашенные участки.
Эритробласт—клетка, имеющая величину от 14 до 18 р.. Форма эритробласта круглая, реже овальная. Ядро располагается в центре нитки, круглое, большое и более компактное, чем у проэритробласта. Хроматин ядра имеет грубую зернистость; у отдельных клеток намечается радиальное расположение базихроматина и выражены тёмные пятна. Нуклеоли в ядре слабо заметны или их нет совсем. Встречаются фигуры митоза с зубчатыми и звёздчатыми краями ядра.
Протоплазма мелкозернистая, синего цвета (базофильиая). У клеток с относительно зрелым ядром протоплазма полихроматофильна, серовато-синего цвета, без зеленого оттенка.
Полихроматофильные эритробласты сравнительно легко отличаются от проэритробластоп. Они меньше размером (10—12—15 [»), имеют грубую структуру ядра без нуклеолей, полпхроматофильную протоплазму в виде узкой и светлой перинуклеарной зоны, окружающей ядро клетки.
Н о р м о б л а с т. Величина у различных животных от 7 до 12 р.. Клетка имеет круглую форму. Ядро у более молодых клеток расположено чаще в центре клетки, реже эксцентрично. Оно сравнительно большое, компактное, темиоокрашиватощееся и без нуклеолей. Расположение хроматина неравномерное. Хроматин имеет вид компактных глыбок с выступающими и советами и трещинами. Рисунок ядра своеобразный и напоминает спицы колеса.
У созревающих нормобластов ядро небольшое, компактное, в сое тояниипикноза—без рисунка. В отдельных клетках ядро распадается на зёрна.
Протоплазма нормобласта различная, что зависит от возраста клетки. У более молодых нормобластов протоплазма слегка полихроматофильная, а у зрелых—ортохроматическая. Чем интенсивнее протоплазма окрашена в синий цвет, тем она меньше содержит гемоглобина, тем клетка моложе. Чем слабее базофилия, тем старше эритробласт и тем больше он насыщен гемоглобином.
Между типичными клетками, описанными выше, находится много переходных форм. Последние дают возможность с несомненностью установить родственную связь между проэритробластом и нормобластом, несмотря на резкую разницу в величине клеток и в структуре их составных частей.
Клетки эритронитарного ряда, потерявшие ядро, носят название нормсцитов—нормальных зрелых эритроцитов, циркулирующих в крови-млекопитающих.
Эритроциты птиц. Эритроциты птиц, по данным Зайцева, Лебедева, Преображенского, Левкович, Зубиной и др., сохраняют характер истинных клеток. Они состоят из ядра и протоплазмы. Протоплазма их соответствует веществу нормоцитов млекопитающих.
Эритроциты птиц имеют овальную, эллипсовидную I форму, с тонким, иногда заострённым краем. В центре I кровяного тельца имеется ядро, а потому клетка в середине значительно толще и в профиль имеет фигуру веретена.
У голубя иногда встречаются безъядерные эритроциты—микроциты, круглой и овальной формы, и эритроциты с полихроматофильной зернистостью. Иногда встречаются «тени» эритроцитов, с неясными, расплывчатыми контурами ядра и светлоокрашивающейся протоплазмой (Левкович). То же самое Зайцев отмечает у кур, предполагая, что это явление связано с гемолитическим процессом.
В свежей крови отдельные эритроциты птиц имеют, как и у млекопитающих, зеленовато-жёлтый цвет. Это относится только к протоплазме. Ядро эритроцита не содержит гемоглобина и просвечивает в виде бесцветного овального тельца. Ядро эритроцита кур имеет ясно выраженную сетчатую хроматиновую основу (Зайцев).
Рис. 2. Кровь гуся.
При окраске, по Гимза, протоплазма эритроцитов розовая, а ядро фиолетовое с хроматиновой глыбчатостью (Левкович)
Число эритроцитов в 1 мм3 крови птиц меньше подвержено видовым и индивидуальным колебаниям, чем у млекопитающих, и стоит в обратном отношении к величине эритроцитов.
Средний диаметр эритроцита птиц следующий (в микронах):
Вопрос о строении ядерных эритроцитов у птиц, так же как и безъядерных у млекопитающих животных, остается открытым. В живом состоянии протоплазма эритроцитов птиц является гомогенной. Наличие оболочки одними аирами признаётся, другими нет. В последнее время особый интерес вызывав строение края. При действии химических веществ, а также при окраске свежей крови генцианой, здесь, непосредственно под свободной поверхностью выступает система взаимно переплетающих нитей и волокон, охватывающих контур тельца. Это образование получило название краевого
Молодые эритроциты птиц в стадии эритробластов содержат в своей протоплазме типичные хондриозомы, но у зрелых эритроцитов от них, видимо, не остается и следа.
Ядро зрелых эритроцитов имеет компактное строение и состоит из тесно расположенных хромитиновых частиц, иногда сливающихся, по видимому, в сплошную массу.
Эритроциты сельскохозяйственных животных принадлежат к числу клеток, богатых плотными составным частям — 40% на 60%, воды. Строма их содержит гемоглобин, лецитин, холестерин, белки и соли. Анализ показывает, что на 1000 частей сухих веществ приходится: гемоглобина 868-944, белка 51-122, лецетин 7,2-35, холестерина 2,5 части. Эритроциты содержат, кроме того, магнезию, фосфорную кислоту И фермент каталазу. Количество солей калия и натрия в крови у различных животных следующее (в процентах):
В сыворотке крови больше натрия, чем калия; в эритроцитах, наоборот, преобладают ионы калия. Наиболее важной составной частью эритроцитов является гемоглобин. В норме в 100 см3 крови содержится 13—14,0 г гемоглобина. Основная функция гемоглобина в организме— переносить кислород вдыхаемого воздуха. Кислород образует с гемоглобином нестойкое соединение—оксигемо-глобин НЬ02. Один грамм гемоглобина способен связать 1,34 см3 кислорода (02).
Кислород легко отщепляется от оксигемоглобина и прочно связывается тканями организма. Гемоглобин, полученный в кристаллическом виде, состоит из двух компонентов: гемохромогена—пигмента, содержащего железо (4,5%), и глобину—безжелезистого белкового вещества (94%).
Способность гемоглобина связывать кислород объясняется именно тем, что гемохромоген имеет железо. Гемо-хромоген в присутствии кислорода переходит в окисленную форму—тематик. Кристаллы гемоглобина у большинства животных принадлежат к ромбической системе, и только у белки кристаллизуются гексагональными табличками. Кристаллы гемоглобина обладают двойным преломлением, пленохроматичны и отклоняют вправо поляризованные лучи.
Из дериватов гемоглобина следует отметить метгемоглобин, который, в отличие от оксигемоглобина, более прочно связывает 02 и с трудом отдаёт его окружающим тканям. Возникая под действием кровяных ядов, а также при отравлении бертолетовой солью и анилином, метге-моглобин понижает дыхательную функцию крови, обусловливая «удушение» тканей—аноксимию. Цвет метгемоглобина красно-бурый (цвет сеции). При разрушений мзтгемоглобина, так же как и оксигемоглобина, образуется гематин. Из других дериватов гемоглобина, при инфекциях и отравлениях некоторыми ядами, обнаруживаются сульфгемоглобин и гематопорфирин.
Каждый дрнь в организме животного разрушается значительное количество эритроцитов. Часть свободного гемоглобина разрушенных эритроцитов перерабатывается в пигмент, содержащий железо, большая же часть перерабатывается в печени до жёлчных пигментов. При усилении распада эритроцитов и функциональной недостаточности печени может появиться желтуха. Если печень не в состоянии утилизировать весь гемоглобин, развивается гемоглобинурия. В настоящее время образование жёлчных пигментов из гемоглобина приписывается ретикуло-эндотелиальной системе селезёнки и печени.
Непрерывный расход гемоглобина пополняется в процессе питания. Накопление и усвоение железа, идущего на построение гемоглобина крови, происходит, повидимому, в костном мозгу, печени и селезёнке.
Красные кровяные тельца млекопитающих животных представляют собой клетки, лишённые ядра и не способные к размножению. Основная физиологическая роль их в организме—разносить кислород тканям и клеткам и выводить отработанный продукт—углекислоту. (На 100 см3 крови приходится 2,5 см3 С02.)
Кругооборот этих двух газов в крови сводится к следующему: в капиллярах гемоглобин легко отдаёт нестойко химически связанный с ним кислород крови тканям, в которых давление 02 к этому времени доходит до 0. Из тканей поступает в кровь накопленная в ней углекислота, способствуя, в свою очередь, отдаче оксигемо-глобином 02 окружающим тканям. Углекислота в дальнейшем легко вытесняется из крови, а гемоглобин вновь насыщается кислородом в альвеолах лёгких.
Ядерные эритроциты птиц и амфибий, по некоторым данным, обладают, по сравнению с безъядерными зрелыми эритроцитами млекопитающих, значительно большей способностью к поглощению кислорода. Способность эритроцитов поглощать 02 удаётся установить аппаратом Баркрофта.
Эритроциты играют известную роль в явлениях иммунитета, Различные яды адсорбируются на эритроцитах, которые затем подвергаются фагоцитозу клетками РЭС.
Эритроциты содержат агглюгиногены (антигены) А и В, благодаря чему они агглютинируются соотв тствующими сыворотками, содержащими агглютинины (антитела) аир.
По данным Збарского и его учеников (Михлин, Зубкова, Мухамедова и др.), эритроциты принимают участие в адсорбции аминокислот, поступающих через капилляры стенок кишечника в кровяное, русло, причём аминокислоты не только пгр»носятся эритроцитами, но эритроциты являются также регуляторами их содержания в плазме крови, поддерживая их концентрацию на более или менее постоянном уровне.
Концентрация аминокислот в эритроцитах может быть подвержена изменениям, но в нормальных условиях она всегда больше единицы (в среднем 1,3—2).
Способность эритроцитов воспринимать аминокислоты падает в редких случаях, например, при уремии, скорбуте, при вытеснении аминокислот камфорой, хинином и хлороформом.
Кроме аминокислот, эритроциты крови способны связывать полипептиды, общий остаточный азот, креатин и креатинин, белки в крови новорождённых, а также переносить адреналин, гистамин, алкалоиды, дифтерийный и столбнячный токсины и некоторые другие вещества.
Таким образом, эритроциты не только принимают участие в процессе гликолиза, но и обладают собственным обменом.
- Подробности
- Раздел: ГЕМАТОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Клеточное ядро — Википедия
У этого термина существуют и другие значения, см. Ядро. Клетки HeLa, ДНК окрашена синим красителем Hoechst[en]. Центральная и правая клетка находятся в интерфазе, и у них окрашено всё ядро. Левая клетка претерпевает митоз, поэтому ДНК конденсирована и окрашено не всё ядро
Клетки HeLa, ДНК окрашена синим красителем Hoechst[en]. Центральная и правая клетка находятся в интерфазе, и у них окрашено всё ядро. Левая клетка претерпевает митоз, поэтому ДНК конденсирована и окрашено не всё ядроКле́точное ядро́ (лат. nucleus) — окружённый двумя мембранами компартмент эукариотической клетки[1] (в клетках прокариот ядро отсутствует). Обычно в клетках эукариот имеется одно ядро, однако некоторые типы клеток, например, эритроциты млекопитающих, не имеют ядра, а другие содержат несколько ядер.
В ядре заключена бо́льшая часть генетического материала клетки, представленного несколькими линейными длинными молекулами ДНК, связанного с белками — хромосомами. Гены, локализованные в хромосомах, составляют ядерный геном. Ядро поддерживает целостность генов, а входящие в его состав белки регулируют клеточные процессы посредством управления экспрессией генов, поэтому ядро является, по сути, контролирующим центром клетки. К основным структурам, из которых состоит ядро, относят ядерную оболочку — двойную мембрану, окружающую ядро и изолирующую его от цитоплазмы, а также ядерный матрикс (который включает ядерную ламину) — сеть филаментов, которая обеспечивает механическую поддержку ядра, подобно цитоскелету в цитоплазме.
Поскольку ядерная оболочка непроницаема для крупных молекул, для регуляции транспорта молекул через ядерную оболочку (ядерный транспорт[en]) служат ядерные поры. Поры пронизывают обе ядерные мембраны и формируют сквозной канал, через который малые молекулы и ионы проходят свободно, а крупные молекулы активно транспортируются с участием белков-переносчиков. Перенос через ядерную оболочку таких крупных молекул, как белки и РНК, необходим для экспрессии генов и поддержания хромосом. Хотя внутри ядра нет окружённых мембраной субкомпартментов, его внутреннее содержимое неоднородно и содержит ряд ядерных телец, которые состоят из особых белков, молекул РНК и частей хромосом. Самое известное ядерное тельце — ядрышко, в котором происходит сборка рибосомных субъединиц. После образования в ядрышке рибосомные субъединицы транспортируются в цитоплазму, где они осуществляют трансляцию мРНК.

Ядро стало первой из органелл, открытых учёными-естествоиспытателями в составе клетки. Самые ранние рисунки клеток и их ядер принадлежат основоположнику научной микроскопии Антони ван Левенгуку (1633—1723), который наблюдал ядро в эритроцитах лосося[2]. Описания ядра также выполнил Франц Бауэр[en] в 1802 году[3], а более детальное описание было выполнено в 1831 году шотландским ботаником Робертом Броуном и представлено на собрании Лондонского Линнеевского общества. Броун изучал орхидеи под микроскопом и обнаружил в клетках наружного слоя цветка непрозрачные области, которые он называл «ареолами» или «ядрами»[4].
Броун не делал предположений относительно функций ядра. В 1838 году Маттиас Шлейден предположил, что ядро участвует в образовании новых клеток, поэтому он ввёл для обозначения ядер термин «цитобласт» (клеточный строитель). Он был уверен, что наблюдал сборку новых клеток вокруг «цитобластов». Убеждённым оппонентом этого взгляда был Франц Мейен, открывший, что клетки размножаются посредством деления, и считавший, что у многих клеток может не быть ядра. Идея об образовании клеток de novo[en], то есть с нуля, посредством цитобластов или иначе, противоречила работам Роберта Ремака (1852) и Рудольфа Вирхова (1855), которые окончательно утвердили новую парадигму, утверждающую, что клетки могут образовываться только из клеток («Omnis cellula e cellula»). Функции ядра оставались неясными[5].
Между 1877 и 1878 годами Оскар Гертвиг опубликовал несколько работ по оплодотворению яиц у морских ежей, в которых показал, что при оплодотворении ядро сперматозоида проникает внутрь яйцеклетки и сливается с её ядром. Впервые было показано, что новая особь развивается из единственной клетки, имеющей ядро. Это противоречило теории Эрнста Геккеля, согласно которой в ходе эмбрионального развития особи последовательно проходятся все этапы филогении её вида, а потому, в частности, поколение первых клеток с ядром якобы образуется из «монерулы» — бесструктурной массы первичной слизи. В связи с этим необходимость ядра сперматозоида для оплодотворения некоторое время была предметом дискуссий. Однако Гертвиг подтвердил свои наблюдения исследованиями на других животных, включая земноводных и моллюсков. В 1884 году Эдуард Страсбургер показал то же самое для растений. Это проложило путь к гипотезе о том, что ядро передаёт наследственный материал. В 1873 году Август Вейсман высказал идею о равнозначности материнского и отцовского материала для наследственности. Функция ядра как носителя генетической информации стала очевидной лишь позже, после открытия митоза и открытия заново законов Менделя в начале XX столетия. На основании этих открытий была сформулирована хромосомная теория наследственности[5].
Ядро — крупнейшая органелла животных клеток[6]. У млекопитающих диаметр ядра составляет примерно 6 мкм, а само ядро составляет около 10 % объёма клетки[7]. Вязкая жидкость, заполняющая ядро, называется нуклеоплазмой и по химическому составу близка к цитозолю, окружающему ядро[8].
Ядерная оболочка и ядерные поры[править | править код]
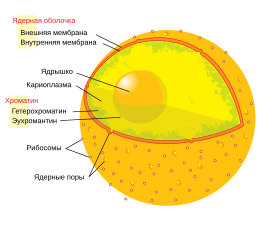 Строение клеточного ядра Строение клеточного ядра |
Ядерная оболочка состоит из двух мембран (наружной и внутренней), которые расположены параллельно на расстоянии от 10 до 50 нм. Ядерная оболочка полностью окружает ядро, отделяя генетический материал клетки от цитоплазмы и служа барьером, предотвращающим свободную диффузию макромолекул между нуклеоплазмой и цитоплазмой. Наружная ядерная мембрана продолжается в мембрану шероховатого эндоплазматического ретикулума (ЭПР) и покрыта рибосомами. Промежуток между ядерными мембранами называется перинуклеарным пространством и продолжается в люмен ЭПР[9].
Ядерные поры, представляющие собой заполненные водой каналы в ядерной оболочке[1], состоят из множества белков, называемых нуклеопоринами. У человека масса пор составляет около 120 000 кДа,что в 40 раз больше массы рибосомы[10]; при этом у дрожжей в состав ядерных пор входит около 50 белков, а у позвоночных — несколько сотен[6]. Хотя диаметр пор составляет 100 нм, ширина щели, через которую могут проходить молекулы, из-за наличия внутри пор регуляторных систем составляет всего 9 нм. В такую щель могут проходить водорастворимые малые молекулы, но не крупные молекулы — такие, как нуклеиновые кислоты и большие белки; для переноса этих молекул в ядро необходим активный (то есть энергозатратный) транспорт. На оболочке ядра типичной клетки млекопитающего располагается от 3000 до 4000 пор[11], и у каждой в месте слияния двух ядерных мембран находится кольцевая структура, имеющая 8 осей симметрии[12]. К кольцу прикрепляется особая структура, известная как ядерная корзина, которая выдаётся в нуклеоплазму, а несколько её филаментов выдаются в цитоплазму. Обе структуры необходимы для опосредования связывания транспортных ядерных белков[6].
Большинство белков, субъединицы рибосом и некоторые ДНК переносятся через ядерные поры посредством семейства транспортных факторов, известных как кариоферины[en]. Кариоферины, опосредующие транспорт в ядро, также называются импортинами[en], а опосредующие транспорт из ядра — экспортинами. Большинство кариоферинов непосредственно взаимодействуют со своим грузом, но некоторые используют для этого адаптерные[en] белки[13]. Стероидные гормоны (такие, как кортизол и альдостерон), а также другие жирорастворимые малые молекулы могут диффундировать в цитоплазму внутрь клетки через клеточную мембрану; в цитоплазме они связываются с белковыми ядерными рецепторами, которые доставляют их в ядро. Здесь ядерные рецепторы[en], связанные со своими лигандами, функционируют как транскрипционные факторы, а в отсутствие лиганда многие рецепторы функционируют как гистондеацетилазы, подавляющие экспрессию некоторых генов[6].
Ядерная ламина[править | править код]
 Строение ядерной оболочки и ядерной ламины
Строение ядерной оболочки и ядерной ламиныВ клетках животных механическую поддержку ядра обеспечивают две сети из промежуточных филаментов: ядерная ламина, представляющая собой сеть промежуточных филаментов на внутренней поверхности ядра, а также менее организованные филаменты на цитозольной поверхности ядра. Обе системы филаментов обеспечивают поддержку ядра и служат для закрепления хромосом и ядерных пор[7].
Ядерная ламина состоит в основном из белков, известных как ламины. Как и все белки, ламины синтезируются в цитоплазме и далее транспортируются внутрь ядра, где они вставляются в ядерную ламину[14][15]. Расположенные на наружной стороне ядерной оболочки ламины (такие, как эмерин[en] и несприн[en]), связываются с элементами цитоскелета, что обеспечивает структурную поддержку ядру. Ламины также обнаруживаются в нуклеоплазме, где они образуют другую регулярную структуру, известную как нуклеоплазматическая вуаль (англ. nucleoplasmic veil)[16]; последнюю можно визуализировать с использованием флуоресцентной микроскопии. Функция вуали неизвестна, но известно, что её нет в ядрышке и она присутствует в интерфазе клеточного цикла[17]. Входящие в состав вуали ламины (такие, как LEM3) связываются с хроматином, и нарушения в их структуре подавляют транскрипцию белоккодирующих генов[18].
Как и другие белки промежуточных филаментов, мономеры ламинов содержат α-спиральный домен, используемый двумя мономерами, чтобы обвиться вокруг друг друга, образуя димер, имеющий структуру биспирали[en]. Два димера далее связываются своими боковыми сторонами в антипараллельной ориентации, образуя тетрамер, известный как протофиламент. Восемь тетрамеров объединяются в скрученный, похожий на верёвку филамент. Филаменты могут собираться и разбираться динамическим образом, то есть длина филамента зависит от относительных скоростей его сборки и разборки[7].
Хромосомы[править | править код]
 Хромосомные территории 24 хромосом человека
Хромосомные территории 24 хромосом человекаВ ядре находится большая часть генетического материала клетки, представленного множеством линейных молекул ДНК, которые организованы в структуры, известные как хромосомы. Суммарная длина молекул ДНК клетки человека составляет около 2 м. В течение большей части клеточного цикла данные молекулы в комплексе с белками формируют так называемый хроматин, а при клеточном делении хромосомы предстают в виде отдельных хорошо различимых хромосом, составляющих кариотип. Небольшое количество клеточного генетического материала располагается в митохондриях и, в случае растительной клетки, в хлоропластах[19].
Известно два вида хроматина. В эухроматине ДНК наименее плотно организована; он содержит гены, которые транскрибируются наиболее часто[19]. Другой вид хроматина, гетерохроматин, более компактен и содержит ДНК, транскрибируемую редко или никогда. Гетерохроматин подразделяется на факультативный, который присутствует только в клетках определённого типа и на определённой стадии клеточного цикла, и конститутивный, представленный такими структурами хромосом, как теломеры и центромеры[20]. В течение интерфазы хроматин каждой хромосомы занимает строго определённую область ядра — хромосомную территорию[21][22]. Активные гены, которые, как правило, располагаются в эухроматине, обычно располагаются на границе хромосомной территории[23].
Ядерные тельца[править | править код]
В ядре клеток млекопитающих содержится ряд дискретных субкомпартментов[24], которые называются ядерными тельцами. Они осуществляют компартментализацию ядра, создавая внутри него отдельные пространства, которые обладают определёнными свойствами. Многие ядерные тельца осуществляют специфические функции — например, синтез и процессинг пре-рибосомных РНК в ядрышке, накопление и сборку компонентов сплайсосом в спеклах (см. ниже) или накопление молекул РНК в параспеклах. Механизмы, которые обеспечивают выполнение ядрышковыми тельцами этих функций, очень разнообразны. В некоторых случаях ядерное тельце может служить местом протекания определённых процессов, например, транскрипции. В других случаях ядерные тельца, по-видимому, опосредованно регулируют локальные концентрации своих компонентов в нуклеоплазме. Подобно цитоплазматическим органеллам, ядерные тельца содержат специфический набор белков, которые определяют их структуру на молекулярном уровне. Однако, в отличие от органелл цитоплазмы, ядерные тельца не окружены липидными мембранами, и их структурная целостность целиком обеспечивается белок-белковыми и РНК-белковыми взаимодействиями. Ниже в таблице перечислены основные характеристики ядерных телец[25].
| Ядерное тельце | Функции | Характерные компоненты | Типичный размер (в мкм) | Количество на ядро |
|---|---|---|---|---|
| Ядрышко | Биогенез рибосом | Машинерия РНК-полимеразы I[en], факторы процессинга рРНК и сборки рибосомных субъединиц | 3—8 | 1—4 |
| Спеклы | Накопление и сборка факторов сплайсинга | Факторы сплайсинга пре-мРНК | 2—3 | 20—50 |
| Стрессовые ядерные тельца | Регуляция транскрипции и сплайсинга в условиях стресса | HSF1[en], HAP | 1—2 | 3—6 |
| Тельце гистоновых локусов | Процессинг пре-мРНК гистонов | NPAT[en], FLASH, U7[en] мяРНП | 0,2—1,2 | 2—4 |
| Тельце Кахаля | Биогенез, созревание и кругооборот малых РНК | Коилин, SMN[en] | 0,2—1,5 | 1—10 |
| PML-тельце | Регуляция стабильности генома, репарация ДНК, контроль транскрипции, защита от вирусов | PML | 0,1—1 | 10—30 |
| Параспеклы | Регуляция мРНК, редактирование РНК | Некодирующие РНК NEAT1/MENε/β, белки PSP1, p54nrb/NONO | 0,2—1 | 2—20 |
| Околоядрышковый компартмент | Посттранскрипционная регуляция набора РНК, синтезированных РНК-полимеразой III[en] | PTB | 0,2—1 | 1—2 |
Ядрышко[править | править код]

Ядрышко — это отдельная плотная структура в ядре. Она не окружена мембраной и формируется в области расположения рДНК — тандемных повторов генов рибосомной РНК (рРНК), называемых ядрышковыми организаторами. Главная функция ядрышка — синтез рРНК и образование рибосом. Структурная целостность ядрышка зависит от его активности, и инактивация генов рРНК приводит к смешению ядрышковых структур[26].
На первой стадии образования рибосом фермент РНК-полимераза I транскрибирует рДНК и образует пре-рРНК, которая далее разрезается на 5,8S, 18S и 28S рРНК[27]. Транскрипция и посттранскрипционный процессинг рРНК происходят в ядрышке при участии малых ядрышковых РНК (snoРНК), некоторые из которых происходят из сплайсированных интронов мРНК генов, кодирующих белки, связанные с работой рибосом. Собранные рибосомные субъединицы — это самые крупные структуры, проходящие через ядерные поры[6].
При рассматривании под электронным микроскопом в ядрышке можно выделить три компонента: фибриллярные центры (ФЦ), окружающий их плотный фибриллярный компонент (ПФК) и гранулярный компонент (ГК), который, в свою очередь, окружает ПФК. Транскрипция рРНК происходит в ФЦ и на границе ФЦ и ПФК, поэтому при активации образования рибосом ФЦ становятся хорошо различимы. Разрезание и модификации рРНК происходят в ПФК, а последующие этапы образования рибосомных субъединиц, включающие загрузку рибосомных белков, происходят в ГК[27].
Тельце Кахаля[править | править код]
 Ядра клеток мыши (синие), содержащие тельца Кахаля (зелёные точки). Изображение получено методом флуоресцентной микроскопии (коилин — маркер телец Кахаля — сращён с зелёным флуоресцентным белком)
Ядра клеток мыши (синие), содержащие тельца Кахаля (зелёные точки). Изображение получено методом флуоресцентной микроскопии (коилин — маркер телец Кахаля — сращён с зелёным флуоресцентным белком)Тельце Кахаля (ТК) — ядерное тельце, имеющееся у всех эукариот. Оно идентифицируется по наличию сигнатурного белка коилина и специфических РНК (scaРНК). В ТК также содержится белок SMN (англ. survival of motor neurons). В ТК наблюдается высокая концентрация сплайсирующих малых ядерных рибонуклеопротеинов (мяРНП) и других факторов процессинга РНК, поэтому считается, что ТК служат местами сборки и/или посттранскрипционной модификации факторов сплайсинга. ТК присутствует в ядре во время интерфазы, но исчезает в митозе. В биогенезе ТК прослеживаются свойства самоорганизующейся структуры[28].
Когда внутриклеточная локализация SMN впервые изучалась методом иммунофлуоресценции, то белок обнаруживался во всей цитоплазме, а также в ядрышковом тельце, сходном по размеру с ТК и часто расположенном рядом с ТК. По этой причине данное тельце было названо «близнецом ТК» (англ. gemini of CB) или просто gem. Однако оказалось, что линия клеток HeLa, в которой было открыто новое тельце, была необычной: в других линиях клеток человека, а также у плодовой мушки Drosophila melanogaster SMN колокализовался с коилином в ТК. Поэтому в общем случае SMN можно рассматривать как важный компонент ТК, а не как маркер отдельного ядерного тельца[29].
Тельце гистоновых локусов[править | править код]
Тельце гистоновых локусов (англ. histone locus body, HLB) содержит факторы, необходимые для процессинга пре-мРНК гистонов. Как и следует из названия, тельца гистоновых локусов ассоциированы с генами, кодирующими гистоны; поэтому предполагается, что в тельцах гистоновых локусов концентрируются факторы сплайсинга. Тельце гистоновых локусов присутствует в клетке во время интерфазы и исчезает с наступлением митоза. Тельце гистоновых локусов нередко рассматривается вместе с тельцем Кахаля по нескольким причинам. Во-первых, в некоторых тельцах гистоновых локусов содержится маркер телец Кахаля — коилин. Во-вторых, эти тельца нередко физически находятся рядом, поэтому между ними наблюдается некоторое взаимодействие. Наконец, очень крупные тельца Кахаля ооцитов земноводных обладают свойствами обоих телец[28].
PML-тельца[править | править код]
Тельца промиелоцитной лейкемии (англ. Promyelocytic leukaemia bodies), или PML-тельца — сферические тельца, разбросанные по всей нуклеоплазме и достигающие около 0,1—1,0 мкм в диаметре. Они известны также под такими названиями, как ядерный домен 10 (англ. nuclear domain 10 (ND10)), тельца Кремера (англ. Kremer bodies) и онкогенные домены PML (англ. PML oncogenic domains). Тельца PML названы по одному из своих ключевых компонентов — белку промиелоцитной лейкемии (PML). Они часто наблюдаются ассоциированными с тельцами Кахаля и тельцами деления (англ. cleavage body)[30]. PML-тельца принадлежат ядерному матриксу и могут быть задействованы в таких процессах, как репликация ДНК, транскрипция и эпигенетический сайленсинг генов[31]. Ключевым фактором организации этих телец выступает белок PML, который привлекает другие белки; последние, по современным представлениям, объединены лишь тем, что они SUMOилированы[en]. Мыши, у которых ген PML делетирован, лишены PML-телец, однако развиваются и живут нормально, поэтому PML-тельца не выполняют незаменимых биологических функций[31].
Спеклы[править | править код]
Спеклы (англ. speckle) — это ядерные тельца, которые содержат факторы сплайсинга пре-мРНК и располагаются в интерхроматиновых участках нуклеоплазмы клеток млекопитающих. При флуоресцентной микроскопии спеклы выглядят как пятнистые тельца неправильной формы, различных размеров, а при электронной микроскопии они выглядят как кластеры интерхроматиновых гранул. Спеклы — динамические структуры, и содержащиеся в них белки и РНК могут перемещаться между спеклами и другими ядерными тельцами, включая участки активной транскрипции. На основании исследований состава, структуры и поведения спеклов была создана модель, объясняющая функциональную компартментализацию ядра и организацию машинерии экспрессии генов[32], сплайсирующих малые ядерные рибонуклеопротеины[33][34] и другие белки, необходимые для сплайсинга пре-мРНК[32]. Из-за изменяющихся потребностей клетки состав и расположение спеклов изменяется согласно транскрипции мРНК и посредством регуляции фосфорилирования специфических белков[35]. Сплайсирующие спеклы также известны как ядерные спеклы, компартменты сплайсирующих факторов, кластеры интерхроматиновых гранул и B-снурпосомы (англ. B snurposomes)[36]. B-снурпосомы найдены в ядрах ооцитов земноводных и зародышах плодовой мушки Drosophila melanogaster[37]. На электронных микрофотографиях B-снурпосомы предстают прикреплёнными к тельцам Кахаля или отдельно от них. Кластеры интерхроматиновых гранул служат местами скопления факторов сплайсинга[38].
Параспеклы[править | править код]
 Микрофотография клеток HeLa с меченым белком параспекл PSP1: 1. цитоплазма; 2. ядро; 3. ядрышко; 4. параспеклы
Микрофотография клеток HeLa с меченым белком параспекл PSP1: 1. цитоплазма; 2. ядро; 3. ядрышко; 4. параспеклыПараспеклы — это ядерные тельца неправильной формы, располагающиеся в интерхроматиновом пространстве ядра[39]. Впервые они были описаны у клеток HeLa, у которых имеется 10—30 параспеклов на ядро, но сейчас параспеклы обнаружены во всех первичных клетках человека, в клетках трансформированных линий и на срезах тканей[40]. Своё название они получили из-за своего расположения в ядре — вблизи спеклов[39].
Параспеклы — динамические структуры, которые изменяются в ответ на изменения в метаболической активности клетки. Они зависят от транскрипции[39], и в отсутствие транскрипции, проводимой РНК-полимеразой II, параспеклы исчезают, а все входящие в их состав белки (PSP1, p54nrb, PSP2, CFI(m)68 и PSF) формируют серповидный околоядрышковый кэп. Этот феномен наблюдается в ходе клеточного цикла: параспеклы присутствуют в интерфазе и всех фазах митоза, за исключением телофазы. В ходе телофазы формируются дочерние ядра, и РНК-полимераза II ничего не транскрибирует, поэтому белки параспеклов и формируют околоядрышковый кэп[40]. Параспеклы участвуют в регуляции экспрессии генов, накапливая те РНК, где есть двухцепочечные участки, которые подвергаются редактированию, а именно превращению аденозина в инозин. Благодаря этому механизму параспеклы задействованы в контроле экспрессии генов при дифференцировке, вирусной инфекции и стрессе[41].
Околоядрышковый компартмент[править | править код]
Околоядрышковый компартмент (ОК) — ядерное тельце неправильной формы, которое характеризуется тем, что располагается на периферии ядрышка. Несмотря на физическую связь, эти два компартмента структурно различны. Обычно ОК обнаруживают в клетках злокачественных опухолей[42]. ОК — динамическая структура, и содержит очень много РНК-связывающих белков и РНК-полимеразу III. Структурная стабильность ОК обеспечивается транскрипцией, осуществляемой РНК-полимеразой III, и наличием ключевых белков. Поскольку присутствие ОК обычно связано со злокачественностью и со способностью к метастазированию, их рассматривают как потенциальные маркеры рака и других злокачественных опухолей. Показана ассоциация ОК со специфическими локусами ДНК[43].
Стрессовые ядерные тельца[править | править код]
Стрессовые ядерные тельца формируются в ядре при тепловом шоке. Они образуются при непосредственном взаимодействии транскрипционного фактора теплового шока 1 (HSF1[en]) и перицентрических тандемных повторов в последовательности сателлита III, что соответствует сайтам активной траснкрипции некодирующих транскриптов сателлита III. Распространено мнение, что такие тельца соответствуют очень плотно упакованным формам рибонуклеопротеиновых комплексов. Считается, что в клетках, подвергающихся стрессу, они участвуют в быстрых, временных и глобальных изменениях в экспрессии генов посредством различных механизмов — например, ремоделирования хроматина и захватывания факторов транскрипции и сплайсинга. В клетках, находящихся в нормальных (не стрессовых) условиях, стрессовые ядерные тельца обнаруживаются редко, однако их количество резко увеличивается под действием теплового шока. Стрессовые ядерные тельца найдены только в клетках человека и других приматов[44].
Ядерные тельца-сироты[править | править код]
Ядерные тельца-сироты (англ. orphan nuclear bodies) — нехроматиновые ядерные компартменты, которые исследованы гораздо хуже, чем другие хорошо охарактеризованные структуры ядра. Некоторые из них выступают как места, в которых белки модифицируются белками SUMO и/или происходит протеасомная деградация белков, помеченных убиквитином[45]. Ниже в таблице приведены характеристики известных ядерных телец-сирот[46].
| Ядерное тельце | Описание | Типичный размер (в мкм) | Количество на ядро |
|---|---|---|---|
| Кластосома | Концентрирует протеасомные комплексы 20S и 19S и белки, связанные с убиквитином. Обнаруживается, главным образом, тогда, когда стимулируется активность протеасом, и разбирается при ингибировании активности протеасом. | 0,2—1,2 | 0—3 |
| Тельце деления (англ. cleavage body) | Обогащено факторами деления CstF[en] и CPSF[en], а также белком DDX1[en], содержащим DEAD-бокс[en]. Обнаруживается в основном в S-фазе, ингибирование транскрипции на него не влияет. | 0,2—1,0 | 1—4 |
| Домен OPT | Обогащён факторами транскрипции Oct1[en] и PTF. Частично колокализуется с сайтами транскрипции. Обнаруживается в основном в поздней G1-фазе, разбирается при ингибировании транскрипции. | 1,0—1,5 | 1—3 |
| Тельце Polycomb | Обнаруживается в клетках человека и дрозофилы, обогащено белком PcG. У человека накапливает белки RING1, BMI1[en], HPC, может быть связано с околоцентромерным гетерохроматином. | 0,3—1,0 | 12—16 |
| Тельце Sam68 | Накапливает белок Sam68 и схожие с ним белки SLM-1 и SLM-2. Разбирается при ингибировании транскрипции. Вероятно, обогащено РНК. | 0,6—1,0 | 2—5 |
| Тельце SUMO | Обогащено белками SUMO и SUMO-конъюгирующим ферментом Ubc9[en]. Концентрирует транскрипционные факторы pCREB, CBP, c-Jun[en]. | 1—3 | 1—3 |
Ядерная оболочка защищает ДНК клетки и участвует в гораздо более сложной регуляции экспрессии генов по сравнению с прокариотической клеткой. У прокариот транскрипция и трансляция являются сопряжёнными процессами и трансляция мРНК в белок начинается ещё до того, как она будет полностью синтезирована. В клетках эукариот цитоплазма, в которой проходит трансляция, и транскрипция, протекающая в ядре, пространственно разобщены, поэтому возникает необходимость в обеспечении транспорта молекул между ядром и цитоплазмой[47].
Ядерная оболочка даёт ядру возможность контролировать своё содержимое и отделяет его от остальной цитоплазмы. Это имеет важное значение для регуляции процессов, протекающих по обе стороны ядерной оболочки. Когда цитоплазматический процесс должен быть как-то ограничен, то обычно его ключевой участник переносится в ядро, где он взаимодействует с факторами транскрипции и таким образом запускает подавление образования некоторых ферментов, задействованных в цитоплазматическом процессе. Например, такой регуляторный механизм имеется у гликолиза — процесса, в ходе которого клетка извлекает энергию из молекулы глюкозы. Первую реакцию гликолиза осуществляет фермент гексокиназа, преобразуя молекулу глюкозы в глюкозо-6-фосфат. Когда концентрация фруктозо-6-фосфата (вещества, в ходе гликолиза образующегося из глюкозо-6-фосфата) возрастает, регуляторный белок отправляет гексокиназу в ядро[48], где она формирует транскрипционный репрессирующий комплекс, который подавляет экспрессию генов, кодирующих ферменты гликолиза[49].
Чтобы контролировать, какие именно гены транскрибируются, в клетке транскрипционные факторы не имеют физического доступа к ДНК, пока они не будут активированы в ходе определённого сигнального пути. Это предотвращает даже низкую экспрессию неправильных генов. В частности, в случае контролируемых NF-κB генов, которые принимают участие в воспалительном процессе, транскрипция индуцируется под действием сигнального пути, например, начинающегося со связывании сигнальной молекулы TNF-α со своим рецептором на клеточной мембране и в конце концов приводящего к активации фактора транскрипции NF-κB. Сигнал ядерной локализации, имеющийся у NF-κB, позволяет ему проходить в ядро и из него через ядерные поры; в ядре он стимулирует транскрипцию генов-мишеней[7].
Компартментализация предотвращает транскрипцию клеткой несплайсированной мРНК. Эукариотические мРНК содержат интроны, которые должны быть удалены до того, как начнётся трансляция мРНК. Сплайсинг, то есть удаление интронов, протекает в ядре, что предотвращает доступ к пре-мРНК рибосом, находящихся вне ядра. Если бы ядра не было, то рибосомы начинали бы транслировать незрелые мРНК, что привело бы к образованию неправильных белковых продуктов[50].
Поскольку транскрипция протекает в ядре, ядро содержит множество белков, непосредственно участвующих в транскрипции или регулирующих этот процесс. К этим белкам относятся хеликазы, которые расплетают двойную спираль ДНК, облегчая доступ к ней других белков, РНК-полимеразы, которые синтезируют РНК, топоизомеразы, влияющие на топологию ДНК, а также разнообразные факторы транскрипции[51].
Выход из ядра и вход в ядро крупных молекул контролируется ядерными порами. Хотя малые молекулы могут проникать в ядро без всякой регуляции, макромолекулы — такие, как белки и РНК — должны связаться с кариоферинами для транспорта в ядро (импортинами) и из ядра (экспортинами). Белки, которые должны быть транспортированы из цитоплазмы в ядро, содержат особую аминокислотную последовательность, известную как сигнал ядерной локализации, с которой связываются импортины. Аналогичным образом белки, которые должны выйти из ядра, содержат сигнал ядерного экспорта[en], распознаваемый экспортинами. Способность импортинов и экспортинов переносить свой груз регулируется ГТФазами — ферментами, которые гидролизуют ГТФ с высвобождением энергии[13]. Ключевая ГТФаза ядерного транспорта — Ran[en], которая может связываться с ГТФ или ГДФ, в зависимости от своего местонахождения (в ядре или в цитоплазме). В ядре взаимодействие Ran-ГТФ с импортином вызывает конформационные изменения в последнем, так что он отделяется от переносимого груза. Образованный комплекс Ran-ГТФ и импортина транспортируется в цитоплазму, где белок RanBP отделяет Ran-ГТФ от импортина. Отделение от импортина позволяет белку GAP[en] связаться с Ran-ГТФ и катализировать гидролиз ГТФ до ГДФ. Далее комплекс Ran-GDP распознаётся белком NUTF2[en], который возвращает его в нуклеоплазму. В ядре белок GEF
Пожалуйста помогите с тестом по биологии 8 класс
Эритроциты Б. Лимфоциты В. Лейкоциты 4. Терморегуляция и гуморальная регуляция в организме осуществляется с помощью: А. Крови + Б. Лимфы В. Тканевой жидкости 5. Межклеточным веществом крови является: А. Вода Б. Плазма + В. Лимфа 6. Мелкие безъядерные клетки крови двояковогнутой формы: А. Эритроциты + Б. Лейкоциты В. Тромбоциты 7. Гемоглобин в составе эритроцитов легко взаимодействует: А. С кислородом + Б. С азотом В. С водородом 8. Срок жизни эритроцитов составляет: А. 30 дней Б. 100–120 дней + В. 5–7 дней 9. Атомы какого металла входят в состав эритроцитов: А. Меди Б. Цинка В. Железа +, у молюсков медь 10. Бесцветные клетки крови, способные к амебоидному движению сквозь стенки сосудов: А. Эритроциты Б. Лейкоциты + В. Тромбоциты 11. Клетки крови, способные вырабатывать антитела: А. Лейкоциты + Б. Тромбоциты В. Лимфоциты 12. Фагоцитоз осуществляют: А. Лейкоциты Б. Лимфоциты В. Эритроциты 13. Уникальная способность клеток крови к фагоцитозу была открыта русским ученым: A. Николаем Ивановичем Пироговым Б. Иваном Петровичем Павловым B. Ильей Ильичом Мечниковым + 14. Лейкоциты образуются: A. В красном костном мозге +, есть ветви краной и белой крови Б. В желтом костном мозге B. В лимфатических узлах 15. В свертывании крови участвуют: А. Эритроциты Б. Тромбоциты +, эритроциты пропитывают тромб В. Лимфоциты 16. Нерастворимый белок плазмы, образующий тромб: А. Фибриноген + фибриноген переходит в фибрин Б. Протромбин В. Фибрин 17. Иммунитет, возникший после перенесения заболевания, является: A. Естественным Б. Искусственным B. Приобретенным + 18. Сыворотка, вводимая в организм больного для борьбы с инфекцией, содержит: A. Активных возбудителей + Б. Антитела против инфекции + B. Ослабленных возбудителей +, может содержать все трикомпонента. 19. Вакцина представляет собой: A. Активных возбудителей Б. Готовые антитела B. Ослабленных возбудителей+ 20. Первую прививку против оспы осуществил: A. Эдвард Дженнер Б. Луи Пастер B. Илья Ильич Мечников Ещё в 1765 г. врачи Суттон и Фьюстер (Fewster) сообщили лондонскому медицинскому обществу, что оспа у дойных коров, если ею заражается человек, предохраняет его от заболевания натуральной человеческой оспой. Лондонское медицинское общество не согласилось с ними, признало их наблюдение простой случайностью, не заслуживающею дальнейшего исследования. Однако, в 1774 г. английский фермер Джестли успешно привил коровьей оспой свою семью, и то же сделал немецкий учитель Плетт в 1791 г. [7] Независимо от них это обнаружил английский врач и натуралист Дженнер, 21. Группы крови были открыты: A. Паулем Эрлихом Б. Карлом Ландштейнером B. Ильей Ильичом Мечниковым Гру́ппа кро́ви — описание индивидуальных антигенных характеристик эритроцитов, определяемое с помощью методов идентификации специфических групп углеводов и белков, включённых в мембраны эритроцитов животных. википедия У человека открыто несколько систем антигенов, основные из них описаны в этой статье. 22. Белки эритроцитов, определяющие группу крови, называются: A. Агглютинины Б. Антитела B. Агглютиногены + 23. У 15 % людей на Земле: A. Положительный резус-фактор Б. Отрицательный резус-фактор + B. Нейтральный резус-фактор 24. Универсальными реципиентами считаются люди: A. С первой и второй группой крови Б. С третьей группой крови B. С четвертой группой крови+, сейчас переливают только одногрупную кровь